Consider how cooking evolves: It starts in the home and then eventually spreads to restaurants and on to cookbooks, along the way transforming a recipe from oral tradition to commercialized product. In the home, recipes are often transmitted from grandmother to mother, or from father to son, or simply by watching and participating. I’ve seen this in rural Mexico, for instance, when an older daughter teaches her younger sister how to pat tortillas the right way. When societies get richer, you start to see restaurants, a form of specialization like auto mechanics or tailors (see: Adam Smith on the division of labor). Restaurants require that strangers — other cooks — be taught the process. That means simplifying or standardizing ingredients so they’re easier to work with and, in many cases, available year-round. This, of course, means writing down the recipe. Once a dish reaches these commercial milestones, cookbooks will follow.
Yes. Why not? It’s a good paragraph. I like this passage as well, which precedes it:
I recall a trip a few years ago to Dar es Salaam, Tanzania, where I was surprised to find that virtually all restaurants were Chinese or Indian. They were excellent, but still I wanted some local food. In a fit of desperation, I paid the maid to make me a Tanzanian dish in the hotel kitchen, a kind of improvised room service, with a large tip attached. I ended up with a sort of porridge that looked quite simple but tasted delicious. As I was enjoying the meal, it occurred to me that writing down the recipe wouldn’t do much good, as I wouldn’t be able to reproduce it at home. The grain — perhaps a maize flour or millet — was unfamiliar, and the rest of the local ingredients were fresher and more delicious than anything I could easily get my hands on at home in Fairfax, Virginia. A recipe like “cook grain; add water and salt” wouldn’t get me far, not even with Whole Foods at my disposal. Continue reading “Stir-fry Memetics”
This is going to be quick, I hope, and dirty, I’m sure. What I’m up to is taking the first crude steps toward an argument about why putting memes in the head makes culture unintelligible.
Dawkins’ central insight, and the only reason to think about memes at all, is that, properly understood, properly cultural evolution is a regime where the beneficiary of successful cultural change (see my post, Roles in Cultural Interaction) is some kind of cultural entity rather than the organism that exhibits, uses, creates, that cultural entity. Call this the memetic regime. In gene-culture coevolution, by contrast, it is the organism that benefits from successful cultural change.
That is, in the regime of gene-culture coevolution, cultural inheritance is simply mode of behavioral inheritance that is different from, and more rapid, than ‘ordinary’ gene-mediated behavioral inheritance. All of animal culture is inherited in this regime. And this regime remains active in human life as well, though it is swamped by the memetic regime.
The question I’m asking is when, and why, in human prehistory did the memetic regime emerge? Stone tools emerge in the archeological record roughly 2.5 million years ago. Finely crafted hand axes–if that, indeed, is what they are–show up 1.5 million years ago. The shapes of these hand axes are conserved over 100s of thousands of years. They don’t change.
On the one hand, these artifacts indicate a level of craftsmanship beyond that we see in any animal. But they don’t show evidence of rapid and directed change. Do they exist fully within the regime of gene-cultural co-evolution? The question is not, of course, simply about the tools and axes themselves, but about the entire way of life in which they are embedded.
I don’t, of course, know the answer to that question. But if they are pre-memetic, then when did the memetic regime emerge, and why?
One obvious inflection point would be the emergence of language as we know it, which seems to have happened between 200,000 and 50,000 years ago. If that’s when the change happened, why? What is it about language that facilitated that change? Continue reading “The Memetic Changeover: When and Why?”
I’ve been thinking a lot about information recently, mostly as a consequence of reading Dan Dennett on memetics. I’m uncomfortable with his usage, and similar ones, and I can’t quite figure out why. Let me offer two passages, and then some comments.
The first passage is from George Williams, a biologist. It’s in a chapter from a book edited by John Brockman, The Third Culture: Beyond the Scientific Revolution:
Evolutionary biologists have failed to realize that they work with two more or less incommensurable domains: that of information and that of matter. I address this problem in my 1992 book, Natural Selection: Domains, Levels, and Challenges. These two domains will never be brought together in any kind of the sense usually implied by the term “reductionism.” You can speak of galaxies and particles of dust in the same terms, because they both have mass and charge and length and width. You can’t do that with information and matter. Information doesn’t have mass or charge or length in millimeters. Likewise, matter doesn’t have bytes. You can’t measure so much gold in so many bytes. It doesn’t have redundancy, or fidelity, or any of the other descriptors we apply to information. This dearth of shared descriptors makes matter and information two separate domains of existence, which have to be discussed separately, in their own terms.
The gene is a package of information, not an object. The pattern of base pairs in a DNA molecule specifies the gene. But the DNA molecule is the medium, it’s not the message. Maintaining this distinction between the medium and the message is absolutely indispensable to clarity of thought about evolution.
Just the fact that fifteen years ago I started using a computer may have had something to do with my ideas here. The constant process of transferring information from one physical medium to another and then being able to recover that same information in the original medium brings home the separability of information and matter. In biology, when you’re talking about things like genes and genotypes and gene pools, you’re talking about information, not physical objective reality. They’re patterns.
I was also influenced by Dawkins’ “meme” concept, which refers to cultural information that influences people’s behavior. Memes, unlike genes, don’t have a single, archival kind of medium. Consider the book Don Quixote: a stack of paper with ink marks on the pages, but you could put it on a CD or a tape and turn it into sound waves for blind people. No matter what medium it’s in, it’s always the same book, the same information. This is true of everything else in the cultural realm. It can be recorded in many different media, but it’s the same meme no matter what medium it’s recorded in.
It seems to me that that is more or less how the concept of information is used in many discussions. It’s certainly how Dennett tends to use it. Here’s a typical passage (it’s the fifth and last footnote in From Typo to Thinko: When Evolution Graduated to Semantic Norms):
There is considerable debate among memeticists about whether memes should be defined as brain-structures, or as behaviors, or some other presumably well-anchored concreta, but I think the case is still overwhelming for defining memes abstractly, in terms of information worth copying (however embodied) since it is the information that determines how much design work or R and D doesn’t have to be re-done. That is why a wagon with spoked wheels carries the idea of a wagon with spoked wheels as well as any mind or brain could carry it.
Here I can’t help but think that Dennett’s pulling a fast one. Information has somehow become reified in a way that has the happy effect of relieving Dennett of the task of thinking about the actual mechanisms of cultural evolution. That in turn has the unhappy effect of draining his assertion of meaning. In what way does a wagon with spoked wheels carry any idea whatsoever, much less the idea of itself? Continue reading “Culture Memes Information WTF!”
Once again, cultural evolution, and the problem of memes: What are they? Where are they? What do they do? While the general case does interest me, culture is so various that it is impossible to think about it directly. One has to think about specific cases. As details are important, I want to choose a fairly specific case, that of jazz in mid-20th-Century America. I want you to imagine that you’re in a jazz club in, say, Philadelphia, in, say, mid-October of 1952. It’s 1:30 in the morning, and the tune is Charlie Parker’s “Dexterity.” The piano player counts it off–ah one, ah two, one two three four…
But we’re getting ahead of ourselves. We need a little conceptual equipment before considering the example. It’s the conceptual equipment that’s in question. Make no mistake, the concept of memes is conceptual equipment, and it’s confused and confusing.
Roles in Cultural Selection
Genes and phenotypes play certain roles in a more or less standard account of biological evolution. The phenotype interacts with the environment, where it either succeeds or fails at reproduction, depending on the “fit” between its traits and that environment. Where the phenotype is successful at reproduction, it is the genes which are said to carry heredity from one generation to the next.
In one very widespread account genes are said to be replicators. That is to say, replication is the role they play in evolutionary change. Here’s what Peter Godfrey-Smith has to say about that (The Replicator in Retrospect, Biology and Philosophy 15 (2000): 403-423.):
In The Selfish Gene (1976), Richard Dawkins had argued that individual genes must be seen as the units of selection in evolutionary processes within sexual populations. This is primarily because the other possible candidates, notably whole organisms and groups, do not “replicate.” Organisms and groups are ephemeral, like clouds in the sky or dust storms in the desert. Only a replicator, which can figure in selective processes over many generations, can be a unit of selection.
At the same time Dawkins coined the term “meme” to name entities filling the replicator role in cultural evolution. Later on he used the term “vehicle” to designate the entity that interacts with the environment. In biological evolution it is phenotypes that are the vehicles. In cultural evolution, well, that’s a matter of some dispute. And that more general dispute–what are the roles in cultural evolution and what kinds of things occupy them?–is what interests me.
However, I don’t particularly like the term “vehicle.” As Godfrey-Smith has noted, following others, it is a gene-centric term, characterizing what entities do from the so-called “gene’s eye” perspective. I’d prefer a more neutral perspective and so will use a term coined by Richard Hull, “interactor.” Here are definitions as Godfrey-Smith gives them:
Still thinking about Dan Dennett’s conception of memetics. He’s got an article in the Encyclopedia of Evolution (Oxford 2005), “New Replicators, The” that’s worth looking at.
Some bits. From the beginning:
…evolution will occur whenever and wherever three conditions are met: replication, variation (mutation), and differential fitness (competition).
In Darwin’s own terms, if there is “descent [i.e., replication] with modification [variation]” and “a severe struggle for life” [competition], better-equipped descendants will prosper at the expense of their competitors. We know that a single material substrate, DNA (with its surrounding systems of gene expression and development), secures the first two conditions for life on earth; the third condition is secured by the finitude of the planet as well as more directly by uncounted environmental challenges.
The first question, then, is whether or not these conditions are met by human culture. Dennett thinks they are and so do I.
From the end, however:
Do any of these candidates for Darwinian replicator actually fulfill the three requirements in ways that permit evolutionary theory to explain phenomena not already explicable by the methods and theories of the traditional social sciences? Or does this Darwinian perspective provide only a relatively trivial unification?
We do not yet know. But are the prospects for non-triviality good enough to warrant considerable investment of conceptual time and energy? And so
We should also remind ourselves that, just as population genetics is no substitute for ecology—which investigates the complex interactions between phenotypes and environments that ultimate yield the fitness differences presupposed by genetics—no one should anticipate that a new science of memetics would overturn or replace all the existing models and explanations of cultural phenomena developed by the social sciences. It might, however, recast them in significant ways and provoke new inquiries in much the way genetics has inspired a flood of investigations in ecology. Continue reading “More on Dennett on Memes”
I’ve been reading around in Dan Dennett’s papers and found this one, The Cultural Evolution of Words and Other Thinking Tools (Cold Spring Harbor Symp Quant Biol, Vol. LXXIV, August, 2009). To be sure, I disagree with his use of the meme concept. To be sure, his use is pretty standard and Dennett, in the standard way, claims more for it than can be justified by the current state of our knowledge and theorizing, but this paper is excellent despite that problem.
As the title indicates, Dennett focuses his attention on words and does so in a way that usefully brings their mystery, if you will, though mystery is rather low on Dennett’s intellectual agenda.
What then are words? Do they even exist? This might seem to be a fatuous philosophical question, composed as it is of the very items it asks about, but it is, in fact, exactly as serious and contentious as the claim that genes do or do not really exist. Yes, of course, there are sequences of nucleotides on DNA molecules, but does the concept of a gene actually succeed (in any of its rival formulations) in finding a perspicuous rendering of the important patterns amidst all that molecular complexity? If so, there are genes; if not, then genes will in due course get thrown on the trash heap of science along with phlogiston and the ether, no matter how robust and obviously existing they seem to us today.
For what it’s worth, I have it on good authority that there are languages which lack a word corresponding to our concept of word, though they generally have a word roughly corresponding to our concept of utterance (you can find this observation in, e.g., Alfred Lord, The Singer of Tales). That doesn’t bear directly on the point Dennett is making in those words as lacking a word for this is that really existing phenomenon is common enough, but it does indicate that words do have a rather diffuse or abstract character that makes it difficult to understand what they are and how they operate.
A bit later Dennett continues:
A promise or a libel or a poem is identified by the words that compose it, not by the trails of ink or bursts of sound that secure the occurrence of those words. Words themselves have physical “tokens” (composed of uttered or heard phonemes, seen in trails of ink or glass tubes of excited neon or grooves carved in marble), and so do genes, but these tokens are a relatively superficial part or aspect of these remarkable information structures, capable of being replicated, combined into elaborate semantic complexes known as sentences, and capable in turn of provoking cognitive, emotional, and behavioral responses of tremendous power and subtly.
I particularly like his phrase in that first sentence, “the trails of ink or bursts of sound that secure the occurrence of those words.” That secure the occurence, that’s nice. “Anchor” might also work, that anchor the occurence of those words in an utterance or a written text, as though the ink or sound were a tether holding the airy nothings of meaning and syntax to the ground. Continue reading “Dan Dennett on Words in Cultural Evolution”
Last month saw the publication of a paper by James and I (our first paper!) on the so-called ‘nomothetic’ approach to links between language structure and social structure. In it we review the recent trend of using large-scale cross-cultural statistical analyses to find links between cultural traits and social structures (e.g. Lupyan & Dale, 2010). We show that statistical tests can be misleading because of the nature of cultural systems. We also argue that using statistics alone does not provide strong explanatory power. However, they can be a valuable part of a pluralistic approach to problems – especially generating hypotheses and as a catalyst for debate. Other approaches can help support the suggestions made by nomothetic studies, such as experiments and models.
Sean Roberts, & James Winters (2012). Social Structure and Language Structure: the New Nomothetic Approach. Psycology of Language Learning, 16 (2), 89-112 : 10.2478/v10057-012-0008-6
Lupyan G, & Dale R (2010). Language structure is partly determined by social structure. PloS one, 5 (1) PMID: 20098492
A somewhat contentious debate among the behavioural sciences is currently underway concerning Mayr’s division of causal explanations in evolutionary theory. Here I’m going to give you a brief rundown of two papers in particular, before I chip in my two-cents about how other insights from the theoretical literature can inform this debate. It seems the discussion is just getting started with respect to cultural evolution, so it’d be interesting to hear other peoples’ comments from either camp.
Over the years, evolutionary theorists have tried to make logical divisions between the kinds of things we can ask about, with a view to making it clear what exactly scientific studies can tell us. A dominant paradigm dividing two levels of causation for biological features we see in the world is Mayr’s distinction between ultimate and proximate causes. Ultimate causation explains the proliferation of a trait in a population in terms of the evolutionary forces acting on that trait. For example, peahens that prefer peacocks with larger tails (an honest signal of fitness following the handicap principle) will have stronger or more successful offspring, and so this preference proliferates along with larger peacock tails. Proximate causation uses immediate physiological and environmental factors to explain a particular peahen’s penchant for a large-tailed peacock in a mate choice trial, where the signal of the peacock’s large tail elevates the hormone levels in the peahen and copulatory behaviour ensues. Although the behaviour in both of these examples is the same, the levels of explanation are based on different sets of factors.
In Perspectives on Psychological Science last year, a paper by Scott-Phillips, Dickins and West voiced some concerns about these two levels of causation being conflated in the behavioural sciences. In particular, they addressed instances where proximate explanations of traits are being framed as ultimate ones. The paper points specifically to studies of the evolution of cooperation, transmitted culture and epigenetics to illustrate this. Regarding the evolution of cooperation, they point to an instance where ‘strong reciprocity’ (an individual’s propensity to reward cooperative norms and sanction violation of these norms) is purported to be an ultimate explanation of why humans cooperate, rather than a proximate mechanism that enables such cooperation.
Table 1 from Scott-Phillips et al. (2011), highlight added.
Among the examples was the feature of linguistic structure (see table 1 from paper above), where several studies pointed to the cultural transmission process as an ultimate explanation of linguistic structure. They suggest that cultural transmission constitutes a proximate process, because it gives the means by which linguistic structure is expressed – and this is how cultural transmission contributes to what the linguistic structure looks like. One analogy might be that the vibrating of my particular vocal cords is a proximate mechanism giving rise, in part, to how my voice sounds, rather than an ultimate explanation of why I vocalise. Since an ultimate account must suggest how a trait contributes to inclusive fitness in order to explain its prevalence in humans, they uncontroversially venture that the ultimate rationale for the ubiquity of linguistic structure is that it greater enables communication (and therefore increases inclusive fitness by enabling cooperative activity).
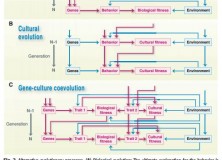
An opposing view was later published in Science by Laland, Sterelny, Odling-Smee et al., who suggest that the use of Mayr’s division of ultimate and proximate causation is not helpful to all evolutionary investigations, and even hampers progress. The grounds for rejecting Mayr’s paradigm seem to lie largely in what Laland et al. term “reciprocal causation”. That is, that “proximate mechanisms both shape and respond to selection, allowing developmental processes to feature in proximate and ultimate explanations”. After aligning proximate explanations with ontogeny and ultimate explanations with phylogeny, they suggest that what we may have called ultimate and proximate features are no longer sharply delineated, and that these reciprocal processes mean that the source of selection sometimes cannot be separated. They present an idea from the field of evolutionary-developmental biology that, if a developmental process makes some variant of a trait more likely to arise than others, then this proximate mechanism helps to construct an “evolutionary pathway”.
Figure 2 from Laland et al. (2011)
The paper also highlights developmental plasticity, and gene-environment interaction more broadly (see fig. 2 from paper, above), as a process where reciprocal causation offers an evolutionary explanation conceptually comparable to ultimate causation. Talking specifically on the topic of linguistic structure, they present the debate about whether specific design features of language are attributable to biological or cultural evolution. The paper points out that cultural evolution determines features of linguistic structure – for example, word order – and that the existing word order determines that of future speakers. Indeed, at the Edinburgh LEC we know that transmission by iterated inductive inference under general conditions can explain particular structures in languages. That cultural evolution determines the variation between languages, Laland et al. say, provides evidence that it is an evolutionary force comparable to natural selection (and, therefore, ultimate explanation).
What follows is a collection of my thoughts on the matter, which are (spoiler alert) largely in support of the Scott-Phillips et al. paper. I hope others more experienced in cultural evolution studies than I will contribute their perspective.
It seems to me that there are a few assumptions made in the Laland et al. paper that are not quite in line with how Mayr himself understood the paradigm. Perhaps much can be learned from this debate’s previous incarnation, when Richard C. Francis made similar arguments against the ultimate/proximate distinction in 1990. In his critique, he equated ultimate causation with phylogeny and proximate causation with ontogeny – an approach that was rebuked by Mayr in 1993, who made the point that “all physiological activities are proximately caused, but is a reflex an ontogenetic phenomenon?”. Mayr’s response is actually rather unhelpful in addressing the arguments fully, and this statement is particularly dense. But what he is getting at here is the idea that interaction with the environment that gives rise to adaptive behaviours (such as recoiling instantly from a hot stove) is itself subject to selection, and thus constitutes a proximate explanation of causation. Relatedly, he points out that most components of the phenotype are indeed the result of genetic contribution and interaction with the environment, which has been successfully explored in biology within the traditional theoretical paradigm.
A perhaps more nuanced account of how we can divide the possible explanations of biological phenomena is offered by Tinbergen in his “four questions”, where ultimate explanations are further subdivided into Function (concerning the adaptive solution to a survival problem favoured by natural selection) and Phylogeny, which is a historical account of when the trait arose in the species, and importantly includes processes other than natural selection that give rise to variation – such as mutation, drift and the constraints imposed by pre-existing traits (see blind spot example below). Proximate explanations are further split into Mechanism (immediate physiological/environmental factors causal in how the trait operates in the individual) and Ontogeny (the way in which this trait develops over the lifetime of the individual). As a simple example, here is the paradigm applied to a trait like mammalian vision that I lifted from Wikipedia: Ultimate Function: To find food and avoid danger. Phylogeny: The vertebrate eye initially developed with a blind spot, but the lack of adaptive intermediate forms prevented the loss of the blind spot. Proximate Causation: The lens of the eye focuses light on the retina Ontogeny: Neurons need the stimulation of light to wire the eye to the brain within a critical period (as those awful studies of blindfolded kittens illustrated).
A schematic below, adapted from Tinbergen (1963) shows how these levels of causation may interact with one another, which appears to communicate something roughly comparable to the importance Laland et al. place on “reciprocal causation” in the formation of adaptive variants:
Adapted from Tinbergen (1963); Causal Relationships
Applied to the debate outlined above, it would seem that there is no apparent reason that a process of gene-environment interaction – including the cultural environment – can’t itself be subject to selection, or that developmental plasticity itself is not an adaptation in need of an ultimate explanation. It has long been the case that behaviour is no longer understood as either “nature” or “nurture”, but gene-environment interaction, with varying levels of heredity. The “reciprocal causation” suggested in Laland et al.’s paper, is (as they point out) very common in nature; feedback loops are uncontroversial proximate processes in biology. That a proximate process may give rise to a dominant variant of a trait in a population does not explain why it is adaptive, and this points to another problem with the proposing the abandonment of Mayr’s paradigm: a logical division of levels of explanation doesn’t seem to be the sort of thing that can be rendered outdated by empirical evidence. Indeed, claims about the particulars of traits and processes (and languages) themselves are a matter for empirical data – but the theoretical issue about the level of explanation that data is useful for does not itself seem to be subject to empirical findings.
The finding that a proximate process such as cultural transmission gives rise to a trait that is prolific in a population is itself exciting and surprising, and even shows us that the pressure for making language easier to learn gives us adaptive languages to learn; however, it could be argued that it is this process that is adaptive, and that the reason why humans so heavily rely on this process is an ultimate explanation.
One way of resolving these two perspectives may be to place cultural processes that give rise to variation at the level of what Tinbergen labels Phylogenetic (one subset of ultimate) explanation, as it concerns processes which produce some heightened frequency of traits over a language’s history. An explanation at the level of Phylogeny still must make recourse to natural selection at some point, since variants that result from mutation or drift are retained because of their adaptive value (or an adaptive trade-off). This approach may be a problem for the current understanding, which holds that the features resulting from cultural processes are themselves adaptive and therefore comparable to what Tinbergen labels Function.
The problem with this is that calling particular structures of language ‘adaptive’ obscures what it is about Language that is actually being selected for. To flesh out what I mean, I think it’s useful to consult Millikan’s (1993) distinction between Direct Proper Function and Derived Proper Function (… bear with me, it’ll be worth it, honest). The Direct Proper Function of a given trait T can be thought of as a “reproduction” of an item that has performed the exact same adaptive function F, and T exists because of these historical performances of F. Sperber and Origgi (2000) use the illustrative example of the heart, where the human heart has a bunch of properties (it pumps blood, makes a thumping noise, etc), but only its ability to pump blood is its Direct Proper Function. This is because even a heart that doesn’t work right or makes irregular thumping noises or whatever, still has the ability to pump blood. Hearts that pump blood have been “reproduced through organisms that, thanks in part to their owning a heart pumping blood, have had descendents similarly endowed with blood-pumping hearts”.
The Derived Proper Function, however, refers to a trait T that is the result of some device that, in some environment, has a Proper Function F. In that given environment, F is usually achieved by the production of something like T. If I unpack this idea and apply it to language, we can understand it as the acquisiton and production of a device that, in this environment, leads to, say, a particular SVO language, T. The Proper Function of adaptive communication is performed by T in this case, but could also be performed by any number of SOV, VSO, etc Ts in other cases. In other words, the Proper Function of this language is not the word order itself, but communication. The word order is the realisation of this device that is reproduced because of the performance of T in a particular environment, but does not necessarily lead to T in the next incarnation of that device (i.e. My child, if born and raised in Japan, will speak Japanese). We see, then, that a proximate process resulting in what a particular language spoken by a given population looks like does not necessarily speak to the evolutionary function. In other words, it is the device that allows the performance of Language that is adaptive, not the individual language itself.
One question being asked in the study of cultural transmission is why a particular language looks like it does, while we also know that there are 6000 different versions that perform the same (ultimate) function. I would even argue that asking how proximate processes shape languages is actually the most exciting and interesting avenue of inquiry precisely because it’s so blindingly obvious what the adaptive function of language is. But perhaps the value in this endeavour is somewhat neglected, in part, because of the same impression that Francis (1990) had: “the attitude, implicit in the term ultimate cause, [is] that these functional analyses are somehow superordinate to those involving proximate causes” which would be a shame. It seems to me that the coarse grain of ultimate vs proximate perhaps doesn’t do enough to help complex proximate study to position itself in the wider theoretical framework, and the best way to proceed from this might be to couch explanation in terms of Function, Phylogeny, Ontogeny and Mechanism. I think more fine-grained terminology grants us more explanatory power, in this case.
A final question in this debate that came up too many times during discussions with the LEC is: what does keeping the traditional paradigm “buy us”? Well, the first answer to this is consilience with one of the most successful and robust theories in science. The same sentiment has been communicated by Pinker and Bloom (1990), who said: “If current theory of language is truly incompatible with the neo-Darwinian theory of evolution, one could hardly blame someone for concluding that it is not the theory of evolution that must be questioned, but the theory of language”. Part of the reason this debate may have arisen is that studies of cultural evolution have used evolutionary theory as an incredibly fruitful way of analysing cultural processes, but additional acknowledgement about how cultural adaptation is different to biological adaptation may be necessary. This difference is an aspect of Laland’s paper (shown in Fig 2) that I think is important, as it’s part of the reason that more nuanced frameworks for cultural evolution are now needed. Without this widespread acknowledgement, cultural evolution may be considered an extension of biological evolutionary theory instead of a successfully applied metaphor. It seems to me that the side of this debate one falls on is well predicted by whether one subscribes to the former interpretation of cultural evolution or the latter.
Knowing which level of explanation current work pertains to is a valuable part of evolutionary exploration, and abandoning this in favour of an approach where proximate processes are explanatory ends to themselves may mean the exploration of Function and Phylogeny may suffer. That said, it is telling, I think, that even in seeking to abandon the proximate/ultimate distinction, we must still exploit this existing terminology in order to explain such a position. That natural selection has explained countless adaptations in all living things is certainly not trivial, and to reject the theory giving rise to ultimate explanations as they’re currently defined is to reject this fundamental aspect of evolutionary theory. The big problem seems to be that we’re coming to understand proximate processes as so elaborate and complex, that a more nuanced framework is needed to deal with the dynamics of those processes. I reckon, however, that such a framework can be developed within the traditional paradigms of evolutionary theory.
References
Francis, R.C. (1990) – “Causes, Proximate and Ultimate” Biology and Philosophy 5(4) 401-415.
Laland, K., Sterelny, K., Odling-Smee, J., Hoppitt, W. & Uller, T. (2011) – “Cause and Effect in Biology Revisited: Is Mayr’s Proximate-Ultimate Distinction Still Useful?” Science 334, 1512-1516.
Mayr, E. (1993) – “Proximate and Ultimate Causations” Biology and Philosophy 8: 93-94.
Millikan, R. (1993) – White Queen Psychology and Other Essays for Alice, Cambridge, Mass: MIT Press.
Pinker, S. & Bloom, P. (1990) – “Natural language and natural selection” Behaviour and Brain Sciences 13, 707-784.
Scott-Phillips, T. Dickins, T. & West, S. (2011) – “Evolutionary Theory and the Ultimate-Proximate Distinction in the Human Behavioural Sciences” Perspectives on Psychological Science 6(1): 38-47.
Sperber, D. & Origgi, G. (2000) – “Evolution, communication and the proper function of language” In P. Carruthers and A. Chamberlain (Eds.) Evolution and the Human Mind: Language, Modularity and Social Cognition (pp.140-169) Cambridge: Cambridge University Press.
Tinbergen, N. (1963) “On Aims and Methods in Ethology,” Zeitschrift für Tierpsychologie, 20: 410–433.
As its name indicates, this post builds on Wild Replicator’s Got Funky Rhythm, Part 1. I want to call your attention, in particular, to the next to the last section, Becoming Memetic. There I trace, albeit sketchily, the history of Rhythm Changes. The point is that Rhymthm Changes didn’t exist as a memetic entity in 1930, when George Gershwin wrote “I Got Rhythm.” Just when the chord changes had become differentiated from the song itself is not clear. But it had certainly happened, at least in the jazz world, by the mid 1940s. Thus, it is not as though certain patterns are essentially memetic while others are not. It’s a question of how the patterns function in the cultural system.
* * * * *
In the previous post I took a look at Rhythm Changes, a memetic entity that has played an important role in jazz and, in particular, in bebop. FWIW, Rhythm Changes has also been used in the theme song for well-known some well-known cartoons, Woody Woodpecker and The Flintstones. In this post I want to do several things:
consider all the elements of “I Got Rhythm,” rather than just the chord changes,
think briefly about how pools of memetic elements function in defining musical styles, and
look briefly at how the chord changes to Gershwin’s tune became memetically active.
Taken together those discussions flesh out the role of memetic elements in music systems in the large. I conclude by
examining this discussion of memes in music in the context of a recent article by Evelyn Fox Keller and David Harel, Beyond the Gene, and not some broad thematic similarities between their discussion and mine.
I Got Rhythm, Whole
As I’ve indicated, Rhythm Changes is derived from, abstracted from, George Gershwin’s “I Got Rhythm.” Now let’s think about the whole tune, not just its harmonic trajectory, i.e. Rhythm Changes. In addition to that trajectory we also have a specific melody, the lyrics, the rhythmic framework, and the arrangement. The lyrics are optional; the tune can be performed without them, and among jazz musicians that is the typical, if not universal, performance practice. Note, however, that any consideration of the lyrics brings a whole other memetic field into consideration, that of language. Continue reading “Wild Replicator’s Got Funky Rhythm, Part 2”
Now that the replicator meme is out and about I’ve got more to say. I’m going to republish two more posts from my 2010 cultural evolution series. These posts are about music. I have various reasons for using music as my cultural evolution conceptual sandbox. For one thing, it means that I don’t have to contend with semantic meanings arbitrarily associated with bits of music. In music, all we’ve got is the physical signal.
In these two posts I choose, not a simple musical example but, rather, a complex one, something jazz musicians know as Rhythm Changes. While I could talk about the four-note motif Beethoven used to construct the first movement of his Fifth Symphony, which is a memetic favorite, that’s too easy. Thinking about it won’t stretch our intuitions about the memetic properties of mere physical things. That motif has four notes, with specific durations and specific note-to-note pitch relationships.
Rhythm Changes isn’t like that. It’s an abstract property of a sound stream. There is now specific number of notes, no specific durations, and no specific note-to-note pitch relationships. Thousands upon thousands of specific musical streams, many quite different from one another, have exemplified the properties of Rhythm Changes.
In the previous post (in this series) I argued memes, the cultural parallel to the biological gene, are those physical properties of objects, events, and processes that allow different individuals to coordinate their participation in those things. In this view, memes are not physical objects, like genes, that spread through a population. Rather, memes are about sharability; they are physical properties that can easily be identified by human nervous systems and thus be the basis for shared (cultural) activity.
In that post I considered a very basic case, people making noise at regular intervals. In that case we have two memes, period (the interval between “hits”) and phase (the relationship between streams of hits by different individuals). Now I want to consider a considerably more complex case, the entity jazz musicians know as Rhythm Changes. This entity assumes that, for a given performance, period length and phase value are agreed upon. In fact it assumes a lot more. We’re dealing with a whole lot of memes here.
But I don’t want to get hung up in those details. I just want to characterize Rhythm Changes in a reasonable way and explain just why I insist that we regard Rhythm Changes as a structured collection of physical properties that can be ascribed to a stream of sound. While it would be nice to characterize Rhythm Changes using the language of acoustics, it’s not at all clear to me that we’ve got the necessary concepts. In any event, if we do, I don’t know them. Instead, I’ll couch my description in the schematic terms jazz musicians tend to use when talking about their craft; these terms are derived, in part, from descriptive and analytic concepts developed for European art music (i.e. classical music).
I’m going do this in two posts, the first will be confined to Rhythm Changes itself. The second will consider how Rhythm Changes came into being and how it functions in the popular music system. Continue reading “Wild Replicator’s Got Funky Rhythm, Part 1”