I’ve been thinking a lot about information recently, mostly as a consequence of reading Dan Dennett on memetics. I’m uncomfortable with his usage, and similar ones, and I can’t quite figure out why. Let me offer two passages, and then some comments.
The first passage is from George Williams, a biologist. It’s in a chapter from a book edited by John Brockman, The Third Culture: Beyond the Scientific Revolution:
Evolutionary biologists have failed to realize that they work with two more or less incommensurable domains: that of information and that of matter. I address this problem in my 1992 book, Natural Selection: Domains, Levels, and Challenges. These two domains will never be brought together in any kind of the sense usually implied by the term “reductionism.” You can speak of galaxies and particles of dust in the same terms, because they both have mass and charge and length and width. You can’t do that with information and matter. Information doesn’t have mass or charge or length in millimeters. Likewise, matter doesn’t have bytes. You can’t measure so much gold in so many bytes. It doesn’t have redundancy, or fidelity, or any of the other descriptors we apply to information. This dearth of shared descriptors makes matter and information two separate domains of existence, which have to be discussed separately, in their own terms.
The gene is a package of information, not an object. The pattern of base pairs in a DNA molecule specifies the gene. But the DNA molecule is the medium, it’s not the message. Maintaining this distinction between the medium and the message is absolutely indispensable to clarity of thought about evolution.
Just the fact that fifteen years ago I started using a computer may have had something to do with my ideas here. The constant process of transferring information from one physical medium to another and then being able to recover that same information in the original medium brings home the separability of information and matter. In biology, when you’re talking about things like genes and genotypes and gene pools, you’re talking about information, not physical objective reality. They’re patterns.
I was also influenced by Dawkins’ “meme” concept, which refers to cultural information that influences people’s behavior. Memes, unlike genes, don’t have a single, archival kind of medium. Consider the book Don Quixote: a stack of paper with ink marks on the pages, but you could put it on a CD or a tape and turn it into sound waves for blind people. No matter what medium it’s in, it’s always the same book, the same information. This is true of everything else in the cultural realm. It can be recorded in many different media, but it’s the same meme no matter what medium it’s recorded in.
It seems to me that that is more or less how the concept of information is used in many discussions. It’s certainly how Dennett tends to use it. Here’s a typical passage (it’s the fifth and last footnote in From Typo to Thinko: When Evolution Graduated to Semantic Norms):
There is considerable debate among memeticists about whether memes should be defined as brain-structures, or as behaviors, or some other presumably well-anchored concreta, but I think the case is still overwhelming for defining memes abstractly, in terms of information worth copying (however embodied) since it is the information that determines how much design work or R and D doesn’t have to be re-done. That is why a wagon with spoked wheels carries the idea of a wagon with spoked wheels as well as any mind or brain could carry it.
Here I can’t help but think that Dennett’s pulling a fast one. Information has somehow become reified in a way that has the happy effect of relieving Dennett of the task of thinking about the actual mechanisms of cultural evolution. That in turn has the unhappy effect of draining his assertion of meaning. In what way does a wagon with spoked wheels carry any idea whatsoever, much less the idea of itself? Continue reading “Culture Memes Information WTF!”
A forthcoming paper (grateful nod to ICCI) in PNAS from Olivier Mascaro and Gergely Csibra presents a series of experiments investigating the representation of social dominance relations in human infants, and it’s excellent news: we’re special.
Social dominance can be inferred in a couple of ways. Causal cues such as age, physical aggression and size can tell us about the dominance status of an individual quite intuitively, so we can make a sensible decision about whether or not we get into a scrap with them. Another way we can establish this is to look for direct realisations of dominance, such as who gets the banana if two hungry chimps both want it; chances are, little Pan Pipsqueak isn’t going to get a look in. In order to be useful, we also have to use this information to expect certain things from the individuals around us, so those representations have some property of stability across time that allows us to have those expectations. The question being explored in this paper is whether the representations we have are about the relationship between the two agents who want the banana, or the individual properties each of them has.
In a series of experiments using preferential looking time as a dependent measure, human infants (9 and 12 month olds) were exposed to videos of geometric figures exhibiting similar goal-directed behaviour. Then they would watch, say, a dominant triangle picking up the last figurative banana when the nondominant pentagon also wanted it. For expository purposes and posterity’s sake, I have constructed an artist’s impression of a dominant triangle and a subordinate pentagon in MSPaint (below, right):
A dominant triangle and subordinate pentagon (artist’s impression).
I’m not just showing off my extraordinary artistic talent here; the good thing about these agents is that there are none of the cues like size or aggression that can give rise to the assignment of individual dominance properties. The task also doesn’t indicate anything similar; it’s just about who gets the desired object when there’s only one left. In other words, the goal-directed actions of two agents are in opposition. After seeing a triangle beat a pentagon to an object of ‘banana’ status, 12 month olds looked for longer when they were then presented with an incongruent trial where the pentagon gained over the triangle. 9 month olds (understandably?) couldn’t care less. So, on the basis of this social interaction alone, the 12 month olds were able to notice when something unexpected happened.
To rule out the possibility that this was just the result of some simple heuristic such as “when triangle and pentagon are present, triangle gets the object” and make sure the infants really were assigning some dominance, another experiment (with 12 and 15 month olds) showed the same test video of the two agents collecting little objects. This time, however, the preceding video was of the triangle dominating a little walled-in space that the pentagon also wanted to inhabit. The 12 month olds had no idea what was up, but the 15 month olds generalised from the first “get out of my room” interaction to the “I get the last banana” interaction. So, 15 month olds can extract, just from watching a social interaction, the dominance status of agents and can generalise that information to novel situations. So if a 15 month old watches you lose your favourite seat in front of the TV, they’ll also expect you to miss out on the last slice of pizza, because you’re a loser.
What we still don’t know is whether they think your belly is inherently yellow, or if you’re just a pushover when interacting with a particular person. Is it the relationship between the triangle and pentagon that the babies are tracking, or do they just give each agent some sort of dominance score? This was addressed in experiment 4, where infants were presented with two interactions: one between A and B, where A wins, and then another between B and C, where B wins. If the babies are assigning an individual value to each agent, they should have some sort of linear, transitive representation of dominance like A > B > C. If they’re then presented with a novel interaction between A and C, they would have the expectation that A will beat C. So if they stare in surprise at a trial where C wins, we know it’s violated that kind of expectation, and that they’re representing this stuff linearly – I.E. each agent has a dominance value. In contrast, if the infant is tracking the relations between agents, they can’t really have an expectation of what will happen when A and C both want a banana, because they’ve never seen C before. The results find that the infants look preferentially when they get an incongruent trial using agent pairs they have seen before – as we’d expect from the previous experiment. When they’re presented with a new “I get the last banana” interaction between A and C, however, there’s nothing startling about it when C wins – which means their expectations are not based on something like A > B > C.
The only tiny little harrumph I have about this result is that all it does is falsify the linear representation account. Though I think their account is absolutely right, it’d be nice to see something more predictive come out of the relation-representation hypothesis that is a little more falsifiable. But this result is pretty huge, and stands in contrast with what we know about social cognition in other animals like baboons (Cheney et al, 1995; Bergman et al, 2003), lemurs (Maclean et al., 2008) and even pigeons (Lazareva & Wasserman, 2012), who seem to employ this sort of hierarchical, transitive inference when presented with novel interactions. It may also muddy the waters a little when we want to make the appealing claim that, since language surely emerged in order to enable communication as we navigated a social environment, hierarchical social cognition gives rise to the processing of languagey things like hierarchical syntax or our semantic representation (Hamilton, 2005), which can be characterised as hierarchical (e.g. hyperonym > hyponym). If we consider the nature of the human social environment, though, it should seem more intuitive that something more reliable than simple transitive inference is necessary in order to successfully navigate through our interactions. Due to our prolific production of (and reliance on) culture, humans have a much more diverse range of social currencies, which correspond to values for things like money, intelligence, blackmail information, who your friends are, ad infinitum. That means it’s pretty reasonable that our social cognition needs new strategies in order to get by; we have a little more to consider than just who’s big and angry enough to get all the bananas.
References
Bergman, T., Beehner, J., Cheney, D. & Seyfarth, R. (2003) “Hierarchical Classification by Rank and Kinship in Baboons” Science 14(302), 1234-1236.
Cheney, D., Seyfarth, R. & Silk, J. (1995) “The response of female baboons (Papio cynocephalus ursinus) to anomalous social interactions: evidence for causal reasoning?” Journal of Comparative Psychology 109(2), 134-141.
Lazareva, O. & Wasserman, E. (2012) “Transitive inference in pigeons: measuring the associative values of stimulus B and D” Behavioural Process 89(3), 244-255.
Maclean, E., Merritt, D. & Brannon, E.M. (2008) “Social complexity predicts transitive reasoning in prosimian primates” Animal Behaviour 76(2), 479-486.
Mascaro, O. & Csibra, G. (forthcoming) “Representation of stable dominance relations by human infants” Proceedings of the National Academy of Sciences
A somewhat contentious debate among the behavioural sciences is currently underway concerning Mayr’s division of causal explanations in evolutionary theory. Here I’m going to give you a brief rundown of two papers in particular, before I chip in my two-cents about how other insights from the theoretical literature can inform this debate. It seems the discussion is just getting started with respect to cultural evolution, so it’d be interesting to hear other peoples’ comments from either camp.
Over the years, evolutionary theorists have tried to make logical divisions between the kinds of things we can ask about, with a view to making it clear what exactly scientific studies can tell us. A dominant paradigm dividing two levels of causation for biological features we see in the world is Mayr’s distinction between ultimate and proximate causes. Ultimate causation explains the proliferation of a trait in a population in terms of the evolutionary forces acting on that trait. For example, peahens that prefer peacocks with larger tails (an honest signal of fitness following the handicap principle) will have stronger or more successful offspring, and so this preference proliferates along with larger peacock tails. Proximate causation uses immediate physiological and environmental factors to explain a particular peahen’s penchant for a large-tailed peacock in a mate choice trial, where the signal of the peacock’s large tail elevates the hormone levels in the peahen and copulatory behaviour ensues. Although the behaviour in both of these examples is the same, the levels of explanation are based on different sets of factors.
In Perspectives on Psychological Science last year, a paper by Scott-Phillips, Dickins and West voiced some concerns about these two levels of causation being conflated in the behavioural sciences. In particular, they addressed instances where proximate explanations of traits are being framed as ultimate ones. The paper points specifically to studies of the evolution of cooperation, transmitted culture and epigenetics to illustrate this. Regarding the evolution of cooperation, they point to an instance where ‘strong reciprocity’ (an individual’s propensity to reward cooperative norms and sanction violation of these norms) is purported to be an ultimate explanation of why humans cooperate, rather than a proximate mechanism that enables such cooperation.
Table 1 from Scott-Phillips et al. (2011), highlight added.
Among the examples was the feature of linguistic structure (see table 1 from paper above), where several studies pointed to the cultural transmission process as an ultimate explanation of linguistic structure. They suggest that cultural transmission constitutes a proximate process, because it gives the means by which linguistic structure is expressed – and this is how cultural transmission contributes to what the linguistic structure looks like. One analogy might be that the vibrating of my particular vocal cords is a proximate mechanism giving rise, in part, to how my voice sounds, rather than an ultimate explanation of why I vocalise. Since an ultimate account must suggest how a trait contributes to inclusive fitness in order to explain its prevalence in humans, they uncontroversially venture that the ultimate rationale for the ubiquity of linguistic structure is that it greater enables communication (and therefore increases inclusive fitness by enabling cooperative activity).
An opposing view was later published in Science by Laland, Sterelny, Odling-Smee et al., who suggest that the use of Mayr’s division of ultimate and proximate causation is not helpful to all evolutionary investigations, and even hampers progress. The grounds for rejecting Mayr’s paradigm seem to lie largely in what Laland et al. term “reciprocal causation”. That is, that “proximate mechanisms both shape and respond to selection, allowing developmental processes to feature in proximate and ultimate explanations”. After aligning proximate explanations with ontogeny and ultimate explanations with phylogeny, they suggest that what we may have called ultimate and proximate features are no longer sharply delineated, and that these reciprocal processes mean that the source of selection sometimes cannot be separated. They present an idea from the field of evolutionary-developmental biology that, if a developmental process makes some variant of a trait more likely to arise than others, then this proximate mechanism helps to construct an “evolutionary pathway”.
Figure 2 from Laland et al. (2011)
The paper also highlights developmental plasticity, and gene-environment interaction more broadly (see fig. 2 from paper, above), as a process where reciprocal causation offers an evolutionary explanation conceptually comparable to ultimate causation. Talking specifically on the topic of linguistic structure, they present the debate about whether specific design features of language are attributable to biological or cultural evolution. The paper points out that cultural evolution determines features of linguistic structure – for example, word order – and that the existing word order determines that of future speakers. Indeed, at the Edinburgh LEC we know that transmission by iterated inductive inference under general conditions can explain particular structures in languages. That cultural evolution determines the variation between languages, Laland et al. say, provides evidence that it is an evolutionary force comparable to natural selection (and, therefore, ultimate explanation).
What follows is a collection of my thoughts on the matter, which are (spoiler alert) largely in support of the Scott-Phillips et al. paper. I hope others more experienced in cultural evolution studies than I will contribute their perspective.
It seems to me that there are a few assumptions made in the Laland et al. paper that are not quite in line with how Mayr himself understood the paradigm. Perhaps much can be learned from this debate’s previous incarnation, when Richard C. Francis made similar arguments against the ultimate/proximate distinction in 1990. In his critique, he equated ultimate causation with phylogeny and proximate causation with ontogeny – an approach that was rebuked by Mayr in 1993, who made the point that “all physiological activities are proximately caused, but is a reflex an ontogenetic phenomenon?”. Mayr’s response is actually rather unhelpful in addressing the arguments fully, and this statement is particularly dense. But what he is getting at here is the idea that interaction with the environment that gives rise to adaptive behaviours (such as recoiling instantly from a hot stove) is itself subject to selection, and thus constitutes a proximate explanation of causation. Relatedly, he points out that most components of the phenotype are indeed the result of genetic contribution and interaction with the environment, which has been successfully explored in biology within the traditional theoretical paradigm.
A perhaps more nuanced account of how we can divide the possible explanations of biological phenomena is offered by Tinbergen in his “four questions”, where ultimate explanations are further subdivided into Function (concerning the adaptive solution to a survival problem favoured by natural selection) and Phylogeny, which is a historical account of when the trait arose in the species, and importantly includes processes other than natural selection that give rise to variation – such as mutation, drift and the constraints imposed by pre-existing traits (see blind spot example below). Proximate explanations are further split into Mechanism (immediate physiological/environmental factors causal in how the trait operates in the individual) and Ontogeny (the way in which this trait develops over the lifetime of the individual). As a simple example, here is the paradigm applied to a trait like mammalian vision that I lifted from Wikipedia: Ultimate Function: To find food and avoid danger. Phylogeny: The vertebrate eye initially developed with a blind spot, but the lack of adaptive intermediate forms prevented the loss of the blind spot. Proximate Causation: The lens of the eye focuses light on the retina Ontogeny: Neurons need the stimulation of light to wire the eye to the brain within a critical period (as those awful studies of blindfolded kittens illustrated).
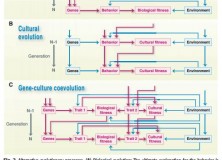
A schematic below, adapted from Tinbergen (1963) shows how these levels of causation may interact with one another, which appears to communicate something roughly comparable to the importance Laland et al. place on “reciprocal causation” in the formation of adaptive variants:
Adapted from Tinbergen (1963); Causal Relationships
Applied to the debate outlined above, it would seem that there is no apparent reason that a process of gene-environment interaction – including the cultural environment – can’t itself be subject to selection, or that developmental plasticity itself is not an adaptation in need of an ultimate explanation. It has long been the case that behaviour is no longer understood as either “nature” or “nurture”, but gene-environment interaction, with varying levels of heredity. The “reciprocal causation” suggested in Laland et al.’s paper, is (as they point out) very common in nature; feedback loops are uncontroversial proximate processes in biology. That a proximate process may give rise to a dominant variant of a trait in a population does not explain why it is adaptive, and this points to another problem with the proposing the abandonment of Mayr’s paradigm: a logical division of levels of explanation doesn’t seem to be the sort of thing that can be rendered outdated by empirical evidence. Indeed, claims about the particulars of traits and processes (and languages) themselves are a matter for empirical data – but the theoretical issue about the level of explanation that data is useful for does not itself seem to be subject to empirical findings.
The finding that a proximate process such as cultural transmission gives rise to a trait that is prolific in a population is itself exciting and surprising, and even shows us that the pressure for making language easier to learn gives us adaptive languages to learn; however, it could be argued that it is this process that is adaptive, and that the reason why humans so heavily rely on this process is an ultimate explanation.
One way of resolving these two perspectives may be to place cultural processes that give rise to variation at the level of what Tinbergen labels Phylogenetic (one subset of ultimate) explanation, as it concerns processes which produce some heightened frequency of traits over a language’s history. An explanation at the level of Phylogeny still must make recourse to natural selection at some point, since variants that result from mutation or drift are retained because of their adaptive value (or an adaptive trade-off). This approach may be a problem for the current understanding, which holds that the features resulting from cultural processes are themselves adaptive and therefore comparable to what Tinbergen labels Function.
The problem with this is that calling particular structures of language ‘adaptive’ obscures what it is about Language that is actually being selected for. To flesh out what I mean, I think it’s useful to consult Millikan’s (1993) distinction between Direct Proper Function and Derived Proper Function (… bear with me, it’ll be worth it, honest). The Direct Proper Function of a given trait T can be thought of as a “reproduction” of an item that has performed the exact same adaptive function F, and T exists because of these historical performances of F. Sperber and Origgi (2000) use the illustrative example of the heart, where the human heart has a bunch of properties (it pumps blood, makes a thumping noise, etc), but only its ability to pump blood is its Direct Proper Function. This is because even a heart that doesn’t work right or makes irregular thumping noises or whatever, still has the ability to pump blood. Hearts that pump blood have been “reproduced through organisms that, thanks in part to their owning a heart pumping blood, have had descendents similarly endowed with blood-pumping hearts”.
The Derived Proper Function, however, refers to a trait T that is the result of some device that, in some environment, has a Proper Function F. In that given environment, F is usually achieved by the production of something like T. If I unpack this idea and apply it to language, we can understand it as the acquisiton and production of a device that, in this environment, leads to, say, a particular SVO language, T. The Proper Function of adaptive communication is performed by T in this case, but could also be performed by any number of SOV, VSO, etc Ts in other cases. In other words, the Proper Function of this language is not the word order itself, but communication. The word order is the realisation of this device that is reproduced because of the performance of T in a particular environment, but does not necessarily lead to T in the next incarnation of that device (i.e. My child, if born and raised in Japan, will speak Japanese). We see, then, that a proximate process resulting in what a particular language spoken by a given population looks like does not necessarily speak to the evolutionary function. In other words, it is the device that allows the performance of Language that is adaptive, not the individual language itself.
One question being asked in the study of cultural transmission is why a particular language looks like it does, while we also know that there are 6000 different versions that perform the same (ultimate) function. I would even argue that asking how proximate processes shape languages is actually the most exciting and interesting avenue of inquiry precisely because it’s so blindingly obvious what the adaptive function of language is. But perhaps the value in this endeavour is somewhat neglected, in part, because of the same impression that Francis (1990) had: “the attitude, implicit in the term ultimate cause, [is] that these functional analyses are somehow superordinate to those involving proximate causes” which would be a shame. It seems to me that the coarse grain of ultimate vs proximate perhaps doesn’t do enough to help complex proximate study to position itself in the wider theoretical framework, and the best way to proceed from this might be to couch explanation in terms of Function, Phylogeny, Ontogeny and Mechanism. I think more fine-grained terminology grants us more explanatory power, in this case.
A final question in this debate that came up too many times during discussions with the LEC is: what does keeping the traditional paradigm “buy us”? Well, the first answer to this is consilience with one of the most successful and robust theories in science. The same sentiment has been communicated by Pinker and Bloom (1990), who said: “If current theory of language is truly incompatible with the neo-Darwinian theory of evolution, one could hardly blame someone for concluding that it is not the theory of evolution that must be questioned, but the theory of language”. Part of the reason this debate may have arisen is that studies of cultural evolution have used evolutionary theory as an incredibly fruitful way of analysing cultural processes, but additional acknowledgement about how cultural adaptation is different to biological adaptation may be necessary. This difference is an aspect of Laland’s paper (shown in Fig 2) that I think is important, as it’s part of the reason that more nuanced frameworks for cultural evolution are now needed. Without this widespread acknowledgement, cultural evolution may be considered an extension of biological evolutionary theory instead of a successfully applied metaphor. It seems to me that the side of this debate one falls on is well predicted by whether one subscribes to the former interpretation of cultural evolution or the latter.
Knowing which level of explanation current work pertains to is a valuable part of evolutionary exploration, and abandoning this in favour of an approach where proximate processes are explanatory ends to themselves may mean the exploration of Function and Phylogeny may suffer. That said, it is telling, I think, that even in seeking to abandon the proximate/ultimate distinction, we must still exploit this existing terminology in order to explain such a position. That natural selection has explained countless adaptations in all living things is certainly not trivial, and to reject the theory giving rise to ultimate explanations as they’re currently defined is to reject this fundamental aspect of evolutionary theory. The big problem seems to be that we’re coming to understand proximate processes as so elaborate and complex, that a more nuanced framework is needed to deal with the dynamics of those processes. I reckon, however, that such a framework can be developed within the traditional paradigms of evolutionary theory.
References
Francis, R.C. (1990) – “Causes, Proximate and Ultimate” Biology and Philosophy 5(4) 401-415.
Laland, K., Sterelny, K., Odling-Smee, J., Hoppitt, W. & Uller, T. (2011) – “Cause and Effect in Biology Revisited: Is Mayr’s Proximate-Ultimate Distinction Still Useful?” Science 334, 1512-1516.
Mayr, E. (1993) – “Proximate and Ultimate Causations” Biology and Philosophy 8: 93-94.
Millikan, R. (1993) – White Queen Psychology and Other Essays for Alice, Cambridge, Mass: MIT Press.
Pinker, S. & Bloom, P. (1990) – “Natural language and natural selection” Behaviour and Brain Sciences 13, 707-784.
Scott-Phillips, T. Dickins, T. & West, S. (2011) – “Evolutionary Theory and the Ultimate-Proximate Distinction in the Human Behavioural Sciences” Perspectives on Psychological Science 6(1): 38-47.
Sperber, D. & Origgi, G. (2000) – “Evolution, communication and the proper function of language” In P. Carruthers and A. Chamberlain (Eds.) Evolution and the Human Mind: Language, Modularity and Social Cognition (pp.140-169) Cambridge: Cambridge University Press.
Tinbergen, N. (1963) “On Aims and Methods in Ethology,” Zeitschrift für Tierpsychologie, 20: 410–433.
The key to the treasure is the treasure.
– John Barth
In view of Sean’s post about Andrew Smith’s take on linguistic replicators I’ve decided to repost this rather longish note from New Savanna. I’d orignally posted it in the Summer of 2010 as part of a run-up to a post on cultural evolution for the National Humanities Center (USA); I’ve collected those notes into a downloadable PDF. Among other things the notes deal with William Croft’s notions (at least as they existed in 2000) and suggests that we’ll find language replicators on the emic side of the emic/etic distinction.
I’ve also appended some remarks I made to John Lawler in the subsequent discussion at New Savanna.
* * * * *
There’s been a fair amount of work done on language from an evolutionary point of view, which is not surprising, as historical linguistics has well-developed treatments of language lineages and taxonomy, the “stuff” of large-scale evolutionary investigation. While this work is directly relevant to a consideration of cultural evolution, however, I will not be reviewing or discussing it. For it doesn’t deal with the theoretical issues that most concern me in these posts, namely, a conceptualization of the genetic and phenotypic entities of culture. This literature is empirically oriented in a way that doesn’t depend on such matters.
The Arbitrariness of the Sign
In particular, I want to deal with the arbitrariness of the sign. Given my approach to memes, that arbitrariness would appear to eliminate the possibility that word meanings could have memetic status. For, as you may recall, I’ve defined memes to be perceptual properties – albeit sometimes very complex and abstract ones – of physical things and events. Memes can be defined over speech sounds, language gestures, or printed words, but not over the meanings of words. Note that by “meaning” I mean the mental or neural event that is the meaning of the word, what Saussure called the signified. I don’t mean the referent of the word, which, in many cases, but by no means all, would have perceptible physical properties. I mean the meaning, the mental event. In this conception, it would seem that that cannot be memetic.
That seems right to me. Language is different from music and drawing and painting and sculpture and dance, it plays a different role in human society and culture. On that basis one would expect it to come out fundamentally different on a memetic analysis.
This, of course, leaves us with a problem. If word meaning is not memetic, then how is it that we can use language to communicate, and very effectively over a wide range of cases? Not only language, of course, but everything that depends on language. Continue reading “In Search of the Wild Replicator”
Replicated Typo 2.0 has reached 100,000 hits! The most popular search term that leads visitors here is ‘What makes humans unique?’ and part of the answer has to be our ability to transmit our culture. But as we’ve shown on this blog, culturally transmitted features can be highly correlated with each other. This fact is a source of both frustration and fascination, so I’ve roped together some of my favourite investigations of cultural correlations into a correlation super-chain. In addition, there’s a whole new spurious correlation at the end of the article!
Digital Humanities Sandbox Goes to the Congo, Part II
While Kurtz is the center of attention in Heart of Darkness, he doesn’t appear until relatively late in the story. He isn’t mentioned until about 8000 words into the 38000 word text nor do we know much about him until a long paragraph that starts roughly 23,000 words into the text. That paragraph, which I’ve called the nexus, is structurally central to the text, and is roughly 1500 words long.
I decided to investigated Kurtz’s presence in the text by the simple expedient of noting where the name “Kurtz” occurs. The result, my colleague Tim Perper subsequently told me, is what’s called a periodogram (PDF):
Figure 1: Periodicity in the appearance of “Kurtz”
Visual inspection suggests that the appearance of “Kurtz” is periodic, with two components, a short one and a significantly longer one. Before discussing this further, however, I would like to explain what I’ve done. Continue reading “Conrad’s Special K: Periodicity in Heart of Darkness”
Prof. Alfred Hubler is an actual mad professor who is a danger to life as we know it. In a talk this evening he went from ball bearings in castor oil to hyper-advanced machine intelligence and from some bits of string to the boundary conditions of the universe. Hubler suggests that he is building a hyper-intelligent computer. However, will hyper-intelligent machines actually give us a better scientific understanding of the universe, or will they just spend their time playing Tetris?
Woah, I just read some of the responses to Dunn et al. (2011) “Evolved structure of language shows lineage-specific trends in word-order universals” (language log here, Replicated Typo coverage here). It’s come in for a lot of flack. One concern raised at the LEC was that, considering an extreme interpretation, there may be no affect of universal biases on language structure. This goes against Generativist approaches, but also the Evolutionary approach adopted by LEC-types. For instance, Kirby, Dowman & Griffiths (2007) suggest that there are weak universal biases which are amplified by culture. But there should be some trace of universality none the less.
Below is the relationship diagram for Indo-European and Uto-Aztecan feature dependencies from Dunn et al.. Bolder lines indicate stronger dependencies. They appear to have different dependencies- only one is shared (Genitive-Noun and Object-Verb).
However, I looked at the median Bayes Factors for each of the possible dependencies (available in the supplementary materials). These are the raw numbers that the above diagrams are based on. If the dependencies’ strength rank in roughly the same order, they will have a high Spearman rank correlation.
Spearman Rank Correlation
Indo-European
Austronesian
Uto-Aztecan
0.39, p = 0.04
0.25, p = 0.19
Indo-European
-0.13, p = 0.49
Spearman rank correlation coefficients and p-values for Bayes Factors for different dependency pairs in different language families. Bantu was excluded because of missing feature data.
Although the Indo-European and Uto-Aztecan families have different strong dependencies, have similar rankings of those dependencies. That is, two features with a weak dependency in an Indo-European language tend to have a weak dependency in Uto-Aztecan language, and the same is true of strong dependencies. The same is true to some degree for Uto-Aztecan and Austronesian languages. This might suggest that there are, in fact, universal weak biases lurking beneath the surface. Lucky for us.
However, this does not hold between Indo-European and Austronesian language families. Actually, I have no idea whether a simple correlation between Bayes Factors makes any sense after hundreds of computer hours of advanced phylogenetic statistics, but the differences may be less striking than the diagram suggests.
UPDATE:
As Simon Greenhill points out below, the statistics are not at all conclusive. However, I’m adding the graphs for all Bayes Factors (these are made directly from the Bayes Factors in the Supplementary Material):
Austronesian: Bantu:
Indo-European: Uto-Aztecan:
Michael Dunn,, Simon J. Greenhill,, Stephen C. Levinson, & & Russell D. Gray (2011). Evolved structure of language shows lineage-specific trends in word-order universals Nature, 473, 79-82
Does the language we speak influence or even shape the way we think? Last December, there was an interesting debate over at The Economist website with Lera Boroditsky defending the motion, and Language Log’s Mark Liberman against the motion (who IMO, both did a very good job).
The result of the online poll was quite clear: 78% agreed with the motion, while 22% disagreed.
There are, however, three main problems with this way of framing the question: First, it’s not really clear what ‘language’ really is, second, the same goes for “thought”, and third, there are many many ways of how “influencing” and “shaping” something can be conceptualized.
In this post I want to focus on the third problem and present a very useful classification system for hypotheses about linguistic relativity outlined in an article by Phillip Wolff and Kevin J. Holmes, which was published in the current issue Wiley Interdisciplinary Review: Cognitive Science.
Just read about an article on phoneme diversity via GNXP and Babel’s Dawn. Hopefully I’ll share some of my thoughts on the paper this weekend as it clearly ties in with work I’m currently doing (see here and here). Below is the abstract:
Human genetic and phenotypic diversity declines with distance from Africa, as predicted by a serial founder effect in which successive population bottlenecks during range expansion progressively reduce diversity, underpinning support for an African origin of modern humans. Recent work suggests that a similar founder effect may operate on human culture and language. here I show that the number of phonemes used in a global sample of 504 languages is also clinal and fits a serial founder-effect model of expansion from an inferred origin in Africa. This result, which is no explained by more recent demographic history, local language diversity, or statistical non-independence within language families, points to parallel mechanisms shaping genetic and linguistic diversity and supports an African origin of modern human languages.
Reference: Atkinson, Q.D (2011). Phonemic Diversity Supports a Serial Founder Effect Model of Language Expansion from Africa. Science 332, 346. DOI: 10.1126/science.1199295.












{kind=link}