The first plenary talk at this year’s Poznań Linguistic Meeting was by Dirk Geeraerts, who is professor of linguistics at the University of Leuven, Belgium.
In his talk, he discussed the possibility that corpus studies could yield evidence against the supposed modularity of language and mind endorsed by, for example, Generative linguists (you can find the abstract here)
Geeraerts began his talk by stating that there seems to be a paradigm shift in linguistics from an analysis of structure that is based on introspection to analyses of behaviour based on quantitative linguistic studies. More and more researchers are adopting quantified corpus-based analyses, which test hypotheses using statistical testing of language behaviour. As a data-set they use experimental data or large corpora. In his talk, he discussed the possibility that corpus studies could yield evidence against the supposed modularity of language and mind endorsed by, for example, Generative linguists (you can find the abstract here)
Multifactoriality
One further trend Geeraerts identified in this paradigm shift is that these kinds of analyses become more and more multifactorial in that they include multiple different factors which are both internal and external to language. Importantly, this way of doing linguistics is fundamentally different than the mainstream late 20th century view of linguistics.
What is important to note here when comparing this trend to other approaches to studying language is that multifactoriality goes against Chomsky’s idea of grammar as an ideal mental system that can be studied through introspection. In the traditional view, it is supposed that there is some kind of ideal language system which everyone has access to. This line of reasoning then justifies introspection as a method of studying the whole system of language and making valid generalizations about it. However, this goes against the emerging corpus linguistic view of language. On this view a random speaker is not representative for the linguistic community as a whole. The linguistic system is not homogenous across all speakers, and therefore introspection doesn’t suffice.
Modularity
The main thrust of Geeraerts’ talk was that research within this emerging paradigm also might call into question the assumption of the modularity of the mind (as advocated, for example by Jerry Fodor or Neil Smith): The view of the mind as a compartmentalized system consisting of discrete components or modules (for example, the visual system, language) plus a central processor.
It’s not about cultural evolution, but I think most people who have even a passing interest in science are gearing up to welcome Higgs Boson to the elementary particle party. Anyway, here’s a nicely put together video on explaining what the Higgs Boson is and why its discovery is significant:
There’s also a more general point about needing to gather a huge amount of data (15 petabytes a year — enough to fill more than 1.7 million dual-layer DVDs a year) to find the very small effect size that is predicted for the Higgs Boson. In itself, data of this magnitude will likely come with significantly more noise, which means physicists have needed to develop well-defined statistical methods (they even have their own statistics committee). It really is a massive achievement for modern science.
The Centre for Philosophy of Science of the Faculty of Science of the Portuguese University of Lisbon is organizing a 3-day international colloquium entitled “From Grooming to Speaking: recent trends in social primatology and human ethology”, on September 10-12th, 2012.
Here’s my opinion in a nutshell: This is a great volume and I’ve really learned a lot from reading it. The authors have done a great job trying to be accessible to an interdisciplinary audience. It’s a great place to start if you’re interested in language evolution or want to get a quick overview of a specific topic in language evolution research. I would’ve liked it if the chapters had a “Further Reading” section, however (like Christiansen and Kirby’s 2003 volume). Some chapters felt a bit too short for me (Steven Mithen‘s chapter on “Musicality and Language” for example is only 3 pages long, Merlin Donald‘s chapter on “the Mimetic Origins of Language” is 4 pages long). I also feel that some topics, like language acquisition, could’ve been dealt with more extensively, but then again, if you compile a handbook, it’s impossible to make everybody happy. Other recent book-length overviews of language evolution (e.g. Fitch’s 2010 book and Hurford’s 2007 and 2012 tomes) are more detailled, but also more technical and not as comprehensive and don’t cover as many topics. To quote my review:
Overall, the Oxford Handbook of Language Evolution is a landmark publication in the field that will serve as a useful guide and reference work through the entanglements and pitfalls of the language evolution jungle for both experienced scholars and newcomers alike.
One last thing I’m particularly unhappy about is that the handbook doesn’t have an Acacia Tree on the cover – which seems like a missed opportunity (kidding).
I’ll try to write about some of my favourite chapters in more detail somewhere down the road/in a couple of weeks.
Gerhard Jäger uses lexostatistics to demonstrate that language similarities can be computed without using tree-based representations (for why this might be important, see Kevin’s post on reconstructing linguistic phylogenies). On the way, he automatically derives a tree of phoneme similarity directly from word lists. The result is an alternative and intuitive look at how languages are related (see graphs below). I review the method, then suggest one way it could get away from prior categorisations entirely.
A few months ago, a documentary I saw on the Discovery Channel covered some research by Graziano Fiorito and colleagues at the Stazione Zoologica in Naples. They were investigating observational learning in wild Octopus vulgaris with a puzzle-box experiment similar to those demonstrating cultural transmission in chimpanzees.
It goes like this: there’s a tasty and terrified crustacean running around in a perspex box that has two possible ways of being opened by hungry octopuses. The experimenters capture a wild octopus (let’s call him Steve) from the harbour (which I’ll get back to in a minute), and they put it in a tank with the puzzle-box. After Steve stares hopelessly at the box for a while, it is then removed from the tank. Steve the kidnapped octopus then gets to watch a captive octopus in the next tank being presented with the same puzzle-box containing the delicious crab. Of course, the captive octopus has been confronted with the puzzle-box enough times that it has worked out a successful solution, and so opens the box like a pro. Steve is then presented with a crab in a puzzle-box again, except this time he goes straight for the crab using the same solution he just learned from the captive octopus. Here is a clip from the documentary, showing a trial of this experiment (NB: not the best quality).
It turns out that the papers on this go back as far as Fiorito & Scotto 1992, and it seems this was the first time observational learning had been demonstrated in invertebrates. The reason I’m interested in reporting this is because the documentary I watched explained another possible motivation/interpretation for Fiorito’s work that I can’t find in any of his actual papers. The octopuses used in the experiments were all caught from the harbour at Naples just before the experiments, which of course controlled for any prior experience with the puzzle boxes. But the results were reported as particularly interesting because the Naples harbour had been overfished and disrupted, resulting in an increase in marine predators that eat the small octopus vulgaris as well as fish that the octopuses themselves rely on. These harsher environmental conditions resulted in the octopuses being forced to inhabit a smaller space alongside each other. As a result, young octopuses were frequently exposed to, and even coexisted with, older octopuses. This is a weird situation for an octopus; they usually live solitary lives and never even meet their own mothers, who die of starvation while caring for the eggs (the fathers die within a few months of having mated). The only real interactions are mating, and conflicts between rival males while competing for a mate.
Toward the end of the documentary, the voiceover growled against some dramatic music about how the combination of observational learning capacities and increased predation pushing octopuses into groups meant that it was only a matter of time before we’re overthrown by octopus vulgaris. This made me think of Dunbar’s “social brain hypothesis” for the emergence of language, and whether I really should prepare to welcome our new octopus overlords. Talking specifically about primates, Dunbar (1996) states that “[primates] in general exhibit two responses to increased predation: they grow physically bigger [or] they increase the size of their groups” (p.110). In order to maintain these groups, that are essential for survival in harsh ecological conditions, Dunbar suggests that standard primate grooming behaviour becomes too time consuming and costly in order to keep up with the rapidly expanding social group, creating a pressure for a more efficient method of bonding and communicating that allows the size of the group to continue increasing. Again with reference just to primate communication, Dunbar says “This [efficient mechanism] need not have involved any dramatic change, for as the studies by Seyfarth and Cheney have shown, primate vocalizations are already capable of conveying a great deal of social information and commentary.” (p.115) In addition to primates, and adding further credence to this idea, it’s been shown that the older Matriarchs of elephant groups make use of vocalisations to seemingly instruct their group on how to fend off lion attacks (McComb et al., 2011; previous Replicated Typo coverage here). It seems to me that the difference between the primates/elephants and the octopuses is that the former endeavour to actually enrich the environment from which their conspecifics extract information; they don’t just learn, they inform. As far as we can tell, there is no such communication like this – that is, enriching the environment in some way that helps other octopuses learn or survive – happening among the octopuses. That said, we know from mating displays and conflicts that cephalopods can communicate with chromatophore signalling. As an interesting aside that is reflective of their cognitive abilities and capacity for suffering, octopuses are treated as honorary vertebrates by UK animal testing laws.
It’s interesting enough that marine biologists (or at least those reporting on marine biologists) seem to have the same idea as Dunbar about the necessary preconditions for successful societies of animals, but why not let’s get wildly speculative? If (..!) existing in groups is in fact adaptive for these octopuses in the face of increased predation, and the competition between them for resources isn’t too great a counter-factor, it seems the only ingredient missing from an octopocalypse is the emergence of some cooperative behaviour. Someone should keep an eye on that harbour.
References
Dunbar, R. (1996) Grooming, Gossip, and the Evolution of Language. Harvard University Press: Cambridge, Massachusetts
Fiorito, G. & Scotto, P. (1992) “Observational learning in Octopus vulgaris” Science 256, 545-546.
McComb, K., Shannon, G., Durant, S., Sayialel, K., Slotow, R., Poole, J. & Moss, C. (2011) “Leadership in elephants: the adaptive value of age” Proceedings of the Royal Society B, published online.
A forthcoming paper (grateful nod to ICCI) in PNAS from Olivier Mascaro and Gergely Csibra presents a series of experiments investigating the representation of social dominance relations in human infants, and it’s excellent news: we’re special.
Social dominance can be inferred in a couple of ways. Causal cues such as age, physical aggression and size can tell us about the dominance status of an individual quite intuitively, so we can make a sensible decision about whether or not we get into a scrap with them. Another way we can establish this is to look for direct realisations of dominance, such as who gets the banana if two hungry chimps both want it; chances are, little Pan Pipsqueak isn’t going to get a look in. In order to be useful, we also have to use this information to expect certain things from the individuals around us, so those representations have some property of stability across time that allows us to have those expectations. The question being explored in this paper is whether the representations we have are about the relationship between the two agents who want the banana, or the individual properties each of them has.
In a series of experiments using preferential looking time as a dependent measure, human infants (9 and 12 month olds) were exposed to videos of geometric figures exhibiting similar goal-directed behaviour. Then they would watch, say, a dominant triangle picking up the last figurative banana when the nondominant pentagon also wanted it. For expository purposes and posterity’s sake, I have constructed an artist’s impression of a dominant triangle and a subordinate pentagon in MSPaint (below, right):
A dominant triangle and subordinate pentagon (artist’s impression).
I’m not just showing off my extraordinary artistic talent here; the good thing about these agents is that there are none of the cues like size or aggression that can give rise to the assignment of individual dominance properties. The task also doesn’t indicate anything similar; it’s just about who gets the desired object when there’s only one left. In other words, the goal-directed actions of two agents are in opposition. After seeing a triangle beat a pentagon to an object of ‘banana’ status, 12 month olds looked for longer when they were then presented with an incongruent trial where the pentagon gained over the triangle. 9 month olds (understandably?) couldn’t care less. So, on the basis of this social interaction alone, the 12 month olds were able to notice when something unexpected happened.
To rule out the possibility that this was just the result of some simple heuristic such as “when triangle and pentagon are present, triangle gets the object” and make sure the infants really were assigning some dominance, another experiment (with 12 and 15 month olds) showed the same test video of the two agents collecting little objects. This time, however, the preceding video was of the triangle dominating a little walled-in space that the pentagon also wanted to inhabit. The 12 month olds had no idea what was up, but the 15 month olds generalised from the first “get out of my room” interaction to the “I get the last banana” interaction. So, 15 month olds can extract, just from watching a social interaction, the dominance status of agents and can generalise that information to novel situations. So if a 15 month old watches you lose your favourite seat in front of the TV, they’ll also expect you to miss out on the last slice of pizza, because you’re a loser.
What we still don’t know is whether they think your belly is inherently yellow, or if you’re just a pushover when interacting with a particular person. Is it the relationship between the triangle and pentagon that the babies are tracking, or do they just give each agent some sort of dominance score? This was addressed in experiment 4, where infants were presented with two interactions: one between A and B, where A wins, and then another between B and C, where B wins. If the babies are assigning an individual value to each agent, they should have some sort of linear, transitive representation of dominance like A > B > C. If they’re then presented with a novel interaction between A and C, they would have the expectation that A will beat C. So if they stare in surprise at a trial where C wins, we know it’s violated that kind of expectation, and that they’re representing this stuff linearly – I.E. each agent has a dominance value. In contrast, if the infant is tracking the relations between agents, they can’t really have an expectation of what will happen when A and C both want a banana, because they’ve never seen C before. The results find that the infants look preferentially when they get an incongruent trial using agent pairs they have seen before – as we’d expect from the previous experiment. When they’re presented with a new “I get the last banana” interaction between A and C, however, there’s nothing startling about it when C wins – which means their expectations are not based on something like A > B > C.
The only tiny little harrumph I have about this result is that all it does is falsify the linear representation account. Though I think their account is absolutely right, it’d be nice to see something more predictive come out of the relation-representation hypothesis that is a little more falsifiable. But this result is pretty huge, and stands in contrast with what we know about social cognition in other animals like baboons (Cheney et al, 1995; Bergman et al, 2003), lemurs (Maclean et al., 2008) and even pigeons (Lazareva & Wasserman, 2012), who seem to employ this sort of hierarchical, transitive inference when presented with novel interactions. It may also muddy the waters a little when we want to make the appealing claim that, since language surely emerged in order to enable communication as we navigated a social environment, hierarchical social cognition gives rise to the processing of languagey things like hierarchical syntax or our semantic representation (Hamilton, 2005), which can be characterised as hierarchical (e.g. hyperonym > hyponym). If we consider the nature of the human social environment, though, it should seem more intuitive that something more reliable than simple transitive inference is necessary in order to successfully navigate through our interactions. Due to our prolific production of (and reliance on) culture, humans have a much more diverse range of social currencies, which correspond to values for things like money, intelligence, blackmail information, who your friends are, ad infinitum. That means it’s pretty reasonable that our social cognition needs new strategies in order to get by; we have a little more to consider than just who’s big and angry enough to get all the bananas.
References
Bergman, T., Beehner, J., Cheney, D. & Seyfarth, R. (2003) “Hierarchical Classification by Rank and Kinship in Baboons” Science 14(302), 1234-1236.
Cheney, D., Seyfarth, R. & Silk, J. (1995) “The response of female baboons (Papio cynocephalus ursinus) to anomalous social interactions: evidence for causal reasoning?” Journal of Comparative Psychology 109(2), 134-141.
Lazareva, O. & Wasserman, E. (2012) “Transitive inference in pigeons: measuring the associative values of stimulus B and D” Behavioural Process 89(3), 244-255.
Maclean, E., Merritt, D. & Brannon, E.M. (2008) “Social complexity predicts transitive reasoning in prosimian primates” Animal Behaviour 76(2), 479-486.
Mascaro, O. & Csibra, G. (forthcoming) “Representation of stable dominance relations by human infants” Proceedings of the National Academy of Sciences
A somewhat contentious debate among the behavioural sciences is currently underway concerning Mayr’s division of causal explanations in evolutionary theory. Here I’m going to give you a brief rundown of two papers in particular, before I chip in my two-cents about how other insights from the theoretical literature can inform this debate. It seems the discussion is just getting started with respect to cultural evolution, so it’d be interesting to hear other peoples’ comments from either camp.
Over the years, evolutionary theorists have tried to make logical divisions between the kinds of things we can ask about, with a view to making it clear what exactly scientific studies can tell us. A dominant paradigm dividing two levels of causation for biological features we see in the world is Mayr’s distinction between ultimate and proximate causes. Ultimate causation explains the proliferation of a trait in a population in terms of the evolutionary forces acting on that trait. For example, peahens that prefer peacocks with larger tails (an honest signal of fitness following the handicap principle) will have stronger or more successful offspring, and so this preference proliferates along with larger peacock tails. Proximate causation uses immediate physiological and environmental factors to explain a particular peahen’s penchant for a large-tailed peacock in a mate choice trial, where the signal of the peacock’s large tail elevates the hormone levels in the peahen and copulatory behaviour ensues. Although the behaviour in both of these examples is the same, the levels of explanation are based on different sets of factors.
In Perspectives on Psychological Science last year, a paper by Scott-Phillips, Dickins and West voiced some concerns about these two levels of causation being conflated in the behavioural sciences. In particular, they addressed instances where proximate explanations of traits are being framed as ultimate ones. The paper points specifically to studies of the evolution of cooperation, transmitted culture and epigenetics to illustrate this. Regarding the evolution of cooperation, they point to an instance where ‘strong reciprocity’ (an individual’s propensity to reward cooperative norms and sanction violation of these norms) is purported to be an ultimate explanation of why humans cooperate, rather than a proximate mechanism that enables such cooperation.
Table 1 from Scott-Phillips et al. (2011), highlight added.
Among the examples was the feature of linguistic structure (see table 1 from paper above), where several studies pointed to the cultural transmission process as an ultimate explanation of linguistic structure. They suggest that cultural transmission constitutes a proximate process, because it gives the means by which linguistic structure is expressed – and this is how cultural transmission contributes to what the linguistic structure looks like. One analogy might be that the vibrating of my particular vocal cords is a proximate mechanism giving rise, in part, to how my voice sounds, rather than an ultimate explanation of why I vocalise. Since an ultimate account must suggest how a trait contributes to inclusive fitness in order to explain its prevalence in humans, they uncontroversially venture that the ultimate rationale for the ubiquity of linguistic structure is that it greater enables communication (and therefore increases inclusive fitness by enabling cooperative activity).
An opposing view was later published in Science by Laland, Sterelny, Odling-Smee et al., who suggest that the use of Mayr’s division of ultimate and proximate causation is not helpful to all evolutionary investigations, and even hampers progress. The grounds for rejecting Mayr’s paradigm seem to lie largely in what Laland et al. term “reciprocal causation”. That is, that “proximate mechanisms both shape and respond to selection, allowing developmental processes to feature in proximate and ultimate explanations”. After aligning proximate explanations with ontogeny and ultimate explanations with phylogeny, they suggest that what we may have called ultimate and proximate features are no longer sharply delineated, and that these reciprocal processes mean that the source of selection sometimes cannot be separated. They present an idea from the field of evolutionary-developmental biology that, if a developmental process makes some variant of a trait more likely to arise than others, then this proximate mechanism helps to construct an “evolutionary pathway”.
Figure 2 from Laland et al. (2011)
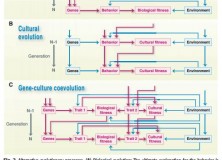
The paper also highlights developmental plasticity, and gene-environment interaction more broadly (see fig. 2 from paper, above), as a process where reciprocal causation offers an evolutionary explanation conceptually comparable to ultimate causation. Talking specifically on the topic of linguistic structure, they present the debate about whether specific design features of language are attributable to biological or cultural evolution. The paper points out that cultural evolution determines features of linguistic structure – for example, word order – and that the existing word order determines that of future speakers. Indeed, at the Edinburgh LEC we know that transmission by iterated inductive inference under general conditions can explain particular structures in languages. That cultural evolution determines the variation between languages, Laland et al. say, provides evidence that it is an evolutionary force comparable to natural selection (and, therefore, ultimate explanation).
What follows is a collection of my thoughts on the matter, which are (spoiler alert) largely in support of the Scott-Phillips et al. paper. I hope others more experienced in cultural evolution studies than I will contribute their perspective.
It seems to me that there are a few assumptions made in the Laland et al. paper that are not quite in line with how Mayr himself understood the paradigm. Perhaps much can be learned from this debate’s previous incarnation, when Richard C. Francis made similar arguments against the ultimate/proximate distinction in 1990. In his critique, he equated ultimate causation with phylogeny and proximate causation with ontogeny – an approach that was rebuked by Mayr in 1993, who made the point that “all physiological activities are proximately caused, but is a reflex an ontogenetic phenomenon?”. Mayr’s response is actually rather unhelpful in addressing the arguments fully, and this statement is particularly dense. But what he is getting at here is the idea that interaction with the environment that gives rise to adaptive behaviours (such as recoiling instantly from a hot stove) is itself subject to selection, and thus constitutes a proximate explanation of causation. Relatedly, he points out that most components of the phenotype are indeed the result of genetic contribution and interaction with the environment, which has been successfully explored in biology within the traditional theoretical paradigm.
A perhaps more nuanced account of how we can divide the possible explanations of biological phenomena is offered by Tinbergen in his “four questions”, where ultimate explanations are further subdivided into Function (concerning the adaptive solution to a survival problem favoured by natural selection) and Phylogeny, which is a historical account of when the trait arose in the species, and importantly includes processes other than natural selection that give rise to variation – such as mutation, drift and the constraints imposed by pre-existing traits (see blind spot example below). Proximate explanations are further split into Mechanism (immediate physiological/environmental factors causal in how the trait operates in the individual) and Ontogeny (the way in which this trait develops over the lifetime of the individual). As a simple example, here is the paradigm applied to a trait like mammalian vision that I lifted from Wikipedia: Ultimate Function: To find food and avoid danger. Phylogeny: The vertebrate eye initially developed with a blind spot, but the lack of adaptive intermediate forms prevented the loss of the blind spot. Proximate Causation: The lens of the eye focuses light on the retina Ontogeny: Neurons need the stimulation of light to wire the eye to the brain within a critical period (as those awful studies of blindfolded kittens illustrated).
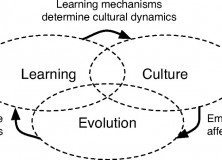
A schematic below, adapted from Tinbergen (1963) shows how these levels of causation may interact with one another, which appears to communicate something roughly comparable to the importance Laland et al. place on “reciprocal causation” in the formation of adaptive variants:
Adapted from Tinbergen (1963); Causal Relationships
Applied to the debate outlined above, it would seem that there is no apparent reason that a process of gene-environment interaction – including the cultural environment – can’t itself be subject to selection, or that developmental plasticity itself is not an adaptation in need of an ultimate explanation. It has long been the case that behaviour is no longer understood as either “nature” or “nurture”, but gene-environment interaction, with varying levels of heredity. The “reciprocal causation” suggested in Laland et al.’s paper, is (as they point out) very common in nature; feedback loops are uncontroversial proximate processes in biology. That a proximate process may give rise to a dominant variant of a trait in a population does not explain why it is adaptive, and this points to another problem with the proposing the abandonment of Mayr’s paradigm: a logical division of levels of explanation doesn’t seem to be the sort of thing that can be rendered outdated by empirical evidence. Indeed, claims about the particulars of traits and processes (and languages) themselves are a matter for empirical data – but the theoretical issue about the level of explanation that data is useful for does not itself seem to be subject to empirical findings.
The finding that a proximate process such as cultural transmission gives rise to a trait that is prolific in a population is itself exciting and surprising, and even shows us that the pressure for making language easier to learn gives us adaptive languages to learn; however, it could be argued that it is this process that is adaptive, and that the reason why humans so heavily rely on this process is an ultimate explanation.
One way of resolving these two perspectives may be to place cultural processes that give rise to variation at the level of what Tinbergen labels Phylogenetic (one subset of ultimate) explanation, as it concerns processes which produce some heightened frequency of traits over a language’s history. An explanation at the level of Phylogeny still must make recourse to natural selection at some point, since variants that result from mutation or drift are retained because of their adaptive value (or an adaptive trade-off). This approach may be a problem for the current understanding, which holds that the features resulting from cultural processes are themselves adaptive and therefore comparable to what Tinbergen labels Function.
The problem with this is that calling particular structures of language ‘adaptive’ obscures what it is about Language that is actually being selected for. To flesh out what I mean, I think it’s useful to consult Millikan’s (1993) distinction between Direct Proper Function and Derived Proper Function (… bear with me, it’ll be worth it, honest). The Direct Proper Function of a given trait T can be thought of as a “reproduction” of an item that has performed the exact same adaptive function F, and T exists because of these historical performances of F. Sperber and Origgi (2000) use the illustrative example of the heart, where the human heart has a bunch of properties (it pumps blood, makes a thumping noise, etc), but only its ability to pump blood is its Direct Proper Function. This is because even a heart that doesn’t work right or makes irregular thumping noises or whatever, still has the ability to pump blood. Hearts that pump blood have been “reproduced through organisms that, thanks in part to their owning a heart pumping blood, have had descendents similarly endowed with blood-pumping hearts”.
The Derived Proper Function, however, refers to a trait T that is the result of some device that, in some environment, has a Proper Function F. In that given environment, F is usually achieved by the production of something like T. If I unpack this idea and apply it to language, we can understand it as the acquisiton and production of a device that, in this environment, leads to, say, a particular SVO language, T. The Proper Function of adaptive communication is performed by T in this case, but could also be performed by any number of SOV, VSO, etc Ts in other cases. In other words, the Proper Function of this language is not the word order itself, but communication. The word order is the realisation of this device that is reproduced because of the performance of T in a particular environment, but does not necessarily lead to T in the next incarnation of that device (i.e. My child, if born and raised in Japan, will speak Japanese). We see, then, that a proximate process resulting in what a particular language spoken by a given population looks like does not necessarily speak to the evolutionary function. In other words, it is the device that allows the performance of Language that is adaptive, not the individual language itself.
One question being asked in the study of cultural transmission is why a particular language looks like it does, while we also know that there are 6000 different versions that perform the same (ultimate) function. I would even argue that asking how proximate processes shape languages is actually the most exciting and interesting avenue of inquiry precisely because it’s so blindingly obvious what the adaptive function of language is. But perhaps the value in this endeavour is somewhat neglected, in part, because of the same impression that Francis (1990) had: “the attitude, implicit in the term ultimate cause, [is] that these functional analyses are somehow superordinate to those involving proximate causes” which would be a shame. It seems to me that the coarse grain of ultimate vs proximate perhaps doesn’t do enough to help complex proximate study to position itself in the wider theoretical framework, and the best way to proceed from this might be to couch explanation in terms of Function, Phylogeny, Ontogeny and Mechanism. I think more fine-grained terminology grants us more explanatory power, in this case.
A final question in this debate that came up too many times during discussions with the LEC is: what does keeping the traditional paradigm “buy us”? Well, the first answer to this is consilience with one of the most successful and robust theories in science. The same sentiment has been communicated by Pinker and Bloom (1990), who said: “If current theory of language is truly incompatible with the neo-Darwinian theory of evolution, one could hardly blame someone for concluding that it is not the theory of evolution that must be questioned, but the theory of language”. Part of the reason this debate may have arisen is that studies of cultural evolution have used evolutionary theory as an incredibly fruitful way of analysing cultural processes, but additional acknowledgement about how cultural adaptation is different to biological adaptation may be necessary. This difference is an aspect of Laland’s paper (shown in Fig 2) that I think is important, as it’s part of the reason that more nuanced frameworks for cultural evolution are now needed. Without this widespread acknowledgement, cultural evolution may be considered an extension of biological evolutionary theory instead of a successfully applied metaphor. It seems to me that the side of this debate one falls on is well predicted by whether one subscribes to the former interpretation of cultural evolution or the latter.
Knowing which level of explanation current work pertains to is a valuable part of evolutionary exploration, and abandoning this in favour of an approach where proximate processes are explanatory ends to themselves may mean the exploration of Function and Phylogeny may suffer. That said, it is telling, I think, that even in seeking to abandon the proximate/ultimate distinction, we must still exploit this existing terminology in order to explain such a position. That natural selection has explained countless adaptations in all living things is certainly not trivial, and to reject the theory giving rise to ultimate explanations as they’re currently defined is to reject this fundamental aspect of evolutionary theory. The big problem seems to be that we’re coming to understand proximate processes as so elaborate and complex, that a more nuanced framework is needed to deal with the dynamics of those processes. I reckon, however, that such a framework can be developed within the traditional paradigms of evolutionary theory.
References
Francis, R.C. (1990) – “Causes, Proximate and Ultimate” Biology and Philosophy 5(4) 401-415.
Laland, K., Sterelny, K., Odling-Smee, J., Hoppitt, W. & Uller, T. (2011) – “Cause and Effect in Biology Revisited: Is Mayr’s Proximate-Ultimate Distinction Still Useful?” Science 334, 1512-1516.
Mayr, E. (1993) – “Proximate and Ultimate Causations” Biology and Philosophy 8: 93-94.
Millikan, R. (1993) – White Queen Psychology and Other Essays for Alice, Cambridge, Mass: MIT Press.
Pinker, S. & Bloom, P. (1990) – “Natural language and natural selection” Behaviour and Brain Sciences 13, 707-784.
Scott-Phillips, T. Dickins, T. & West, S. (2011) – “Evolutionary Theory and the Ultimate-Proximate Distinction in the Human Behavioural Sciences” Perspectives on Psychological Science 6(1): 38-47.
Sperber, D. & Origgi, G. (2000) – “Evolution, communication and the proper function of language” In P. Carruthers and A. Chamberlain (Eds.) Evolution and the Human Mind: Language, Modularity and Social Cognition (pp.140-169) Cambridge: Cambridge University Press.
Tinbergen, N. (1963) “On Aims and Methods in Ethology,” Zeitschrift für Tierpsychologie, 20: 410–433.
Do you have a better memory than a chimp? Tetsuro Matsuzawa demonstrated the amazing working memory abilities of Chimpanzees, but maybe humans can be just as good, with enough practice. Justin Quillinan and I present the Quick-Hold Improvement Challenge (or QHImp Qhallenge). Play our game and find out if you can beat a chimpanzee.
You can see the results update live here. Results so far are tantalizing: Replicated Typo’s very own James Winters has already reached the Ayumu benqhmark (9 numbers viewed for only 209 milliseconds)!
Tetsuro Matsuzawa presented his work on chimpanzees in a plenary talk at Evolang. Matsuzawa covered several very interesting experiments and findings, including an experiment into the working memory of chimpanzees. Ayumu is a chimpanzee who was trained to recognise Arabic numerals on a touch-screen and press them in sequence. The most impressive aspect was that Ayumu could complete the task even when the numbers were only displayed for 210 milliseconds before being masked (the ‘eidetic memory task’ or ‘limited-hold’ memory task, Inoue & Matsuzawa, 2007):
… Except in the Light of Biological and Cultural Evolution
Sean mentioned in one of his many Evolang posts that, based on de Boer’s talk, the real audience for researchers of cultural evolution should be biologists. Well, deciding that actions plus words can work far better together, I decided to get in contact with Jeremy Yoder of the excellent group blog, Nothing in Biology Makes Sense. The result: an introductory post on the biological and cultural evolution of language called Crossing Those Curious Parallels (after Darwin’s famous passage describing the similarities between linguistic and biological change). Most regular readers will be familiar with the content and argument as the article is a pastiche of earlier pieces I wrote on this blog, but there is a sprinkling of some original paragraphs here and there. So feel free to go over, leave a comment and help foster some cross-disciplinary discussions. Actually, on cross-disciplinary note: since physicists seem so keen to solve problems in linguistics, maybe we should lend them a hand and run a corpus analysis to discover that elusive mass of the Higgs boson.