Human speech universally exhibits a 3- to 8-Hz rhythm, corresponding to the rate of syllable production, which is reflected in both the sound envelope and the visual mouth movements. Artificial perturbation of the speech rhythm outside the natural range reduces speech intelligibility, demonstrating a perceptual tuning to this frequency band. One theory posits that the mouth movements at the core of this speech rhythm evolved through modification of ancestral primate facial expressions. Recent evidence shows that one such communicative gesture in macaque monkeys, lip-smacking, has motor parallels with speech in its rhythmicity, its developmental trajectory, and the coordination of vocal tract structures. Whether monkeys also exhibit a perceptual tuning to the natural rhythms of lip-smacking is unknown. To investigate this, we tested rhesus monkeys in a preferential-looking procedure, measuring the time spent looking at each of two side-by-side computer-generated monkey avatars lip-smacking at natural versus sped-up or slowed-down rhythms. Monkeys showed an overall preference for the natural rhythm compared with the perturbed rhythms. This lends behavioral support for the hypothesis that perceptual processes in monkeys are similarly tuned to the natural frequencies of communication signals as they are in humans. Our data provide perceptual evidence for the theory that speech may have evolved from ancestral primate rhythmic facial expressions.
Writing in Nature, last week, Techumseh Fitch wrote a short news article on Ghazanfar’s findings including a very concise but clear outline on the two main hypotheses for the evolutionary origin of human speech, which he also goes over in his 2010 book. Namely, the hypothesis that speech is derived from primate vocalizations as the same vocal production system (lungs, larynx and vocal tract) is used to produce both primate calls and speech. However, as Fitch states, “a problem is that human speech is unique among primate vocalizations in being a very flexible, learned signal, whereas most primate calls have a strong, species-specific genetic determination. The ‘vocal origins’ hypothesis favours evolutionary continuity of vocal production over a hypothetical discontinuity in vocal control and vocal learning.”
The second hypothesis is MacNeilage’s hypothesis that speech rhythms originated not in the vocal, but in the visual domain. As the mouth generates not just vocal, but also visual, signals. The strength in this hypothesis lies in the fact that these articulators are under learned voluntary control in non-human primates. MacNeilage argues that speech develops in babies’ babbling as a lip-smacking behaviour superimposed on a vocal signal. Fitch states: “This rhythmic stream is then differentially modified, by learned tongue and lip movements, into the vowels and consonants of speech. Support for this hypothesis comes from previous work demonstrating that the detailed kinematics of lip-smacking are strikingly similar to those of speech. But Ghazanfar and colleagues’ work adds support from the domain of perception, indicating that perceptual tuning for the two signal classes is also consistent with MacNeilage’s hypothesis.”
As has been covered on this blog before, a lot of research on speech evolution has focused on the descended larynx. This new research adds to the body of work that suggest that anatomy might not be as important as first imagined, and that neural control and vocal learning may be much more important.
The 20th anniversary special commemorative issue of Pragmatics & Cognition features a number of interesting articles which comment on linguist Dan Everett‘s 2012 book “Language: The Cultural Tool“. In this book, Everett, who is best known for his work on the indigenous language Pirahã, argues for the important of culture and interaction and against the Chomskyan idea of innately specified, specifically linguistic knowledge or architecture. The issue also includes replies by Dan Everett to each of his commentators.
Unfortunately, the articles are all behind a paywall, but the abstracts already make some interesting points.
“language use, and coordinated communication more generally, is an emergent product of human self-organization processes. Both broad regularities and specific variations in linguistic structure and behavior can be accounted for by self-organizational processes that operate without explicit internal rules, blueprints, or mental representations. A major implication of this view is that both linguistic patterns and behaviors, within and across speakers, emerge from the dynamical interactions of brain, body, and world, which gives rise to highly context-sensitive and varied linguistic performances.”
After Berwick, Friederici, Chomsky, Bolhuis (2013) last month, Berwick has contributed to another paper featuring evidence from birdsong. Miyagawa, Berwick and Okanoya (2013) is published in frontiers of psychology here. Abstract below:
We propose a novel account for the emergence of human language syntax. Like many evolutionary innovations, language arose from the adventitious combination of two pre-existing, simpler systems that had been evolved for other functional tasks. The first system, Type E(xpression), is found in birdsong, where the same song marks territory, mating availability, and similar “expressive” functions. The second system, Type L(exical), has been suggestively found in non-human primate calls and in honeybee waggle dances, where it demarcates predicates with one or more “arguments,” such as combinations of calls in monkeys or compass headings set to sun position in honeybees. We show that human language syntax is composed of two layers that parallel these two independently evolved systems: an “E” layer resembling the Type E system of birdsong and an “L” layer providing words. The existence of the “E” and “L” layers can be confirmed using standard linguistic methodology. Each layer, E and L, when considered separately, is characterizable as a finite state system, as observed in several non-human species. When the two systems are put together they interact, yielding the unbounded, non-finite state, hierarchical structure that serves as the hallmark of full-fledged human language syntax. In this way, we account for the appearance of a novel function, language, within a conventional Darwinian framework, along with its apparently unique emergence in a single species.
This seems to be slightly in contrast to Berwick, Friederici, Chomsky, Bolhuis (2013), which at times seems to hint that looking at abilities in other species is probably useless because language is so special and specific to humans, but it does share a lot of the same themes.
There has been a lot of talk round these parts recently of the merits of pluralistic approaches to problems in language evolution, and condemning the assignment of too much explanatory power to statistical correlations away from other forms of evidence, such as cultural learning experiments. Sean and James recently published a paper about this here which includes some commentary on Hay & Bauer (2007), who find that speaker population size and phoneme inventory size correlate (the more speakers a language has, the bigger its phoneme inventory is). James has blogged about this extensively here. More recently Moran, McCloy & Wright presented a critical analysis of Hay & Bauer’s (2007) findings here along with a statistical analysis of their own which uses more languages than Hay & Bauer (2007), and finds little to no correlation between speaker population and various measures of the phonological system, I hope James will do a blog about this as the resident expert.
As I’ve just mentioned, doing further statistical analysis is one good way of disputing or confirming the results of large scale statistical studies. But turning to experimental evidence is also a good way to back up the findings of statistical results and to tease out patterns of causation. I discuss this briefly here.
Recently, I was reading Selten & Warglien (2007) (mentioned by James here and covered by John Hawks here), which is a study which looks at how simple languages emerge within a coordination task with no initial shared language. The experiment uses pairwise interactions in which participants had to refer to figures which could be distinguished using features on three levels of outer shape, inner shape and colour (see picture). Participants were given a code which had a limited number of letters which they were to use to communicate with one another. However, the use of letters within this code had a cost within the language game the participants were playing, so the less letters they used the higher their score. Also, the more communicatively successful they were, the higher their score.
The study was primarily interested in what enhanced the emergence of structure in this code via the communication game. They looked at the effects of 2 variables, the number of letters available and variability in the set of figures. I am only going to discuss the effects of the first variable here. Selten & Warglien (2007) start off with an experiment where only two (and then three) letters were available which showed very little convergence to a common code. A common code is defined as being a code where the signals for all figures agree between the two participants. However, when given a larger inventory of letters to play with, participants were much more successful at creating a common code. This is not surprising as more symbols permit a higher degree of cost efficiency within the language game as you can use more distinct, shorter expressions. Selten & Warglien (2007) also make the point that the human capability to produce a large variety of phonetic signals seems to be at the root of the emergence of most linguistic structure, because if you only have a small inventory of individual units, you have to rely more on positional structure. Positional systems are systems like the Arabic number notation which are more likely invented rapidly rather than the product of slow emergence via cultural evolution, but can be easily used once they have emerged.
This is all very interesting in its own right, but the reason I brought it up in this post is that Selten & Warglien (2007) have shown that you can experimentally explore the effects of the size of inventory on an artificial language in a laboratory setting. I know that the natural direction of causation is to assume that demographic structure (e.g. the size of a population) affects the linguistic structure (e.g. the size of the phoneme inventory), but it might be possible to see whether a common code can be more easily reached within a small language community using only a small number of phonemes, than with a larger speaker community. I’m also not sure how one might create an experimental proxy for size of population in an experiment such as this (perhaps repeated interaction between the same participants compared with interaction within changing pairs). It might also be possible to look at the effects that the size of inventory can have on other linguistic features that have been hypothesised to correlate with population size, e.g. how regular the compositional structure of an emerging language is given difference inventory sizes.
References
Hay, J., & Bauer, L. (2007). Phoneme inventory size and population size Language, 83 (2), 388-400 DOI: 10.1353/lan.2007.0071
Roberts, S. & Winters, J. (2012). Social Structure and Language Structure: the New Nomothetic Approach. Psychology of Language and Communication, 16(2), pp. 79-183. Retrieved 12 Feb. 2013, from doi:10.2478/v10057-012-0008-6
Selten, R., & Warglien, M. (2007). The emergence of simple languages in an experimental coordination game Proceedings of the National Academy of Sciences, 104 (18), 7361-7366 DOI: 10.1073/pnas.0702077104
There is a “Skeptics In The Pub” event in Glasgow on March 4th, where Dr Thom Scott-Phillips will be discussing the perceptions and misconceptions of evolutionary psychology, in light of the public backlash against it that seems to be increasing all the time. This kind of public engagement is very sorely needed if we are to combat the rampant misinformation that crops up in both academic and non-academic communities. Among the criticisms being addressed at the event are the claims that evolutionary psychology is sexist, racist, or otherwise politically problematic. This is an important discussion.
From what I see around the feminist blogosphere, evolutionary psychology has a bad rap. Some recent examples I’ve come across include comments such as: “This new junk science named “evolutionary psychology” is the last variant of the male supremacy bible, following Freud’s mythology” and “[the way this article approaches the problem] is a bad idea [because] It smacks of evo psyche”. Even more liberal feminist blogs such as The F Word UK toe a similar line: Josephine Tsui seems to be on a personal mission against Evolutionary Psychology, armed with such ludicrous arguments as “You cannot replicate Evolutionary Psychology therefore it fails the methodologies of science” which display both an immature line of thinking and a fundamental misunderstanding of the theoretical motivations and methodologies entailed. Needless to say I’ve never seen this criticism leveraged against Evolutionary Biology, despite it being applicable to both.
Evolutionary psychology has a sound theoretical basis; it has been well established that natural selection is a means by which complex life and complex behaviour occurs. This tends to worry political movements like feminism, which has its roots in social constructionism. Such worry is unfounded; there is certainly a role for social constructionism within an evolutionary account of human behaviour. Put broadly, our plastic brains depend on complex social learning and pedagogy, which is an established cornerstone of human success. This ability to respond to (and be shaped by) the cultural environment has itself been selected for in humans, and can account for all manner of behaviours from language to mating preferences. Keep reading for a demonstration of how evolutionary psychology can in fact lend itself very well to the goal of engineering of social change.
So, on one side of the sexually selective understanding coin is a worried feminist movement, who risk losing a good grasp of evolutionary psychology by dismissing it entirely. On the other side, are the misogynist (mis)interpretations that have inspired this trepidation in the first place. That evolutionary psychology is abused and misinterpreted by misogynists and racists (and let’s be real here, this has happened a lot) is the problem, and it’s a serious one with real political consequences. Just this year, Steve Moxon submitted evidence to parliament (and was subsequently invited to speak) against the development of measures to encourage women in the workplace. Evolutionary psychology formed the backbone of his case, and he is not alone. Only an informed public can approach these claims with adequate discernment, so it is important that we address how some claims are morally wrong and incorrect. But it is also as important to discuss why they do not represent anything inherent about evolutionary psychology as a discipline.
We can illustrate the first way that evolutionary psychology can be wrong by using the problem of eugenics. Eugenics is theoretically sound, in as much as we know that we can selectively breed to a criteria and expect a predictable result; we’ve been doing it with dogs for 10 thousand years. This is also morally wrong and should not be attempted in humans. Just because eugenics is morally reprehensible, however, doesn’t mean we say the principles of artificial (or natural) selection aren’t true. Nor should this be the case for evolutionary psychology as a field; that it has been misapplied/misinterpreted within our social context (or just says something that we don’t like) simply does not speak to how scientifically correct it is. Another way that the interpretation of these studies can be grossly wrong is the Naturalistic Fallacy; the idea that if something is natural, it is inherently good or should be normative. This is obviously untrue; my human body is adapted to long-distance running, but I reject outright the idea that this is something I ought to engage in.
While citing the naturalistic fallacy is a good answer to most any claim about innate human proclivities, I think it’s also necessary to refute specific claims on their own grounds where possible. The final way for evolutionary psychology to be wrong is simply that rationalisation isn’t science, and instances where it is being passed off as such can be exposed for what they are. To illustrate, we generally do not dismiss the entirety of modern medicine as false because of the historical mistreatment of pregnant women in childbirth by doctors. Here, we can see that those occurrences are indeed morally wrong. However, it’s also the case that those instances are bad medicine by medicine’s own standards. Similarly, instances of bad science in evolutionary psychology, where latent misogyny and racism rears its head, can be refuted on their own grounds. This can and should be done without blithely dismissing the entire field.
It is a disaster that large factions of social justice movements are on the verge of outright anti-intellectualism when it comes to evolutionary psychology. Preserving ignorance about the field with out-of-hand dismissal neglects the potential for this tool to contribute to worthwhile political goals. We don’t have to stop at simply refuting the harmful instances of bad evolutionary study; there is also a positive agenda to be highlighted here. In the spirit of this, I’d like to share a preview of some work I’ve been hobbying with Justin Quillinan, inspired by a recent paper called “Asia’s Missing Women: A Problem in Applied Evolutionary Psychology?”. The paper aims to explore sex-preferential parental investment, which is a prolific problem in parts of Asia, the Middle East and North Africa, where the population’s sex-ratios are heavily male skewed as a result. It is already well documented that women suffer like this, so what can an evolutionary analysis can bring to the table? The problem, as presented in the paper, is this:
Asia’s missing women are, in economic terms, an aggregate outcome of millions of parenting decisions. The individual drivers behind those decisions emerge from interactions between our evolved parenting preferences and social and economic circumstances.
How do we untangle this seemingly nebulous problem? How do we determine why many self-perceived individuals act in such a similar way that the net result is the literal eradication of the female class? The approach of the paper seems to be largely influenced by economics, which is fairly central to a lot of evolutionary work. Game theory can be paraphrased as something like “given that the rules of the game are (x, y, z), which strategy should I employ to reap the most advantageous outcome?”. The most successful strategy is the one that is most likely to survive in the population, and hence it is the one we most expect to find. If we treat the problem of parents favouring boys over girls as a solution to the problem of parents’ circumstances, the question then becomes “what are the parameters that make this survival strategy worth employing such that it is so common?”
By comparing the commonalities in cultures that have this problem, Brooks identifies some ecomonic and social factors that may reward preferential parental investment. This is important: it means that campaigners for change don’t have to simply say the reason that girls are selectively aborted or neglected starts and ends with “girls are undervalued in these cultures”. Despite how true that is, it is also true of many cultures who do not have skewed sex ratios, and doesn’t really point to any concrete way of tackling the problem. If we can identify the driving factors that make parents behave this way with an evolutionary analysis, it means we can target specific structures with a specific end goal in mind.
In an interesting and wide-ranging investigation, the paper compares skewed parental investment occurence in non-human animals with the social and historical particulars that have led to this behaviour in disparate human populations. In doing so, Brooks proposes that male sex-biased populations are the systematic result of a population’s patrilineage, patrilocal kinship systems, and the dowry system. It was my hunch that the sex-biased population ratio could be reducible to patrilocality alone; that is, the system whereby women leave their blood relatives in order to live with (and care for) their husband’s family when they marry. Let us assume that the number of blood relatives in your family is a proxy measure of fitness. In a patrilocal social order, it is necessarily the case that having a son is more advantageous than having a daughter – precisely because daughters will always leave. Let us now also factor in the effects of infanticide and abortion; the option to neglect/kill your male/female offspring according to whether or not the most successful families you know had a boy or girl, will lead to the preferential elimination of females.
So we’ve implemented a model (source code available here) demonstrating exactly this:
We start with a population of agents separated into a number of families of a single breeding pair each single individuals. At each time-step, the following events occur: 1. Reproduction: fertile breeding pairs of agents have a new agent ‘child’ of random sex. A fertile pair is one that does not have an unmarried child that is younger than the age of maturity. 1.2. Abortion: Now, the pair can choose to either keep or to abort their new agent. To make this decision, they choose a random family, with larger families having a proportionally higher probability of being chosen. If the sex ratio of that family is a mismatch to the child they have just had, they will abort the child. Otherwise, they will keep it. (ETA: the abortion decision is based on the sex ratio of all the offspring of the chosen family). 2. Marriage: Every single, mature agent attempts to pair with a random, opposite sex partner from a different family, to form a breeding pair. In patrilocality, the female leaves her family and is appended to her husband-agent’s family to form a breeding pair. (In the matrilocality sanity check, the situation is vice versa and male agents join their wives’ families). 3. Death: Agents above a certain age are removed from the population. If a family no longer has any members, we generate a new breeding pair individual so that the population doesn’t die out.
The first null model was as above, with the omission of step 1.2. We later implemented one that is as above, but minus patrilocal marriage (ie, married agents simply form a new family pair) because this is a better comparison.
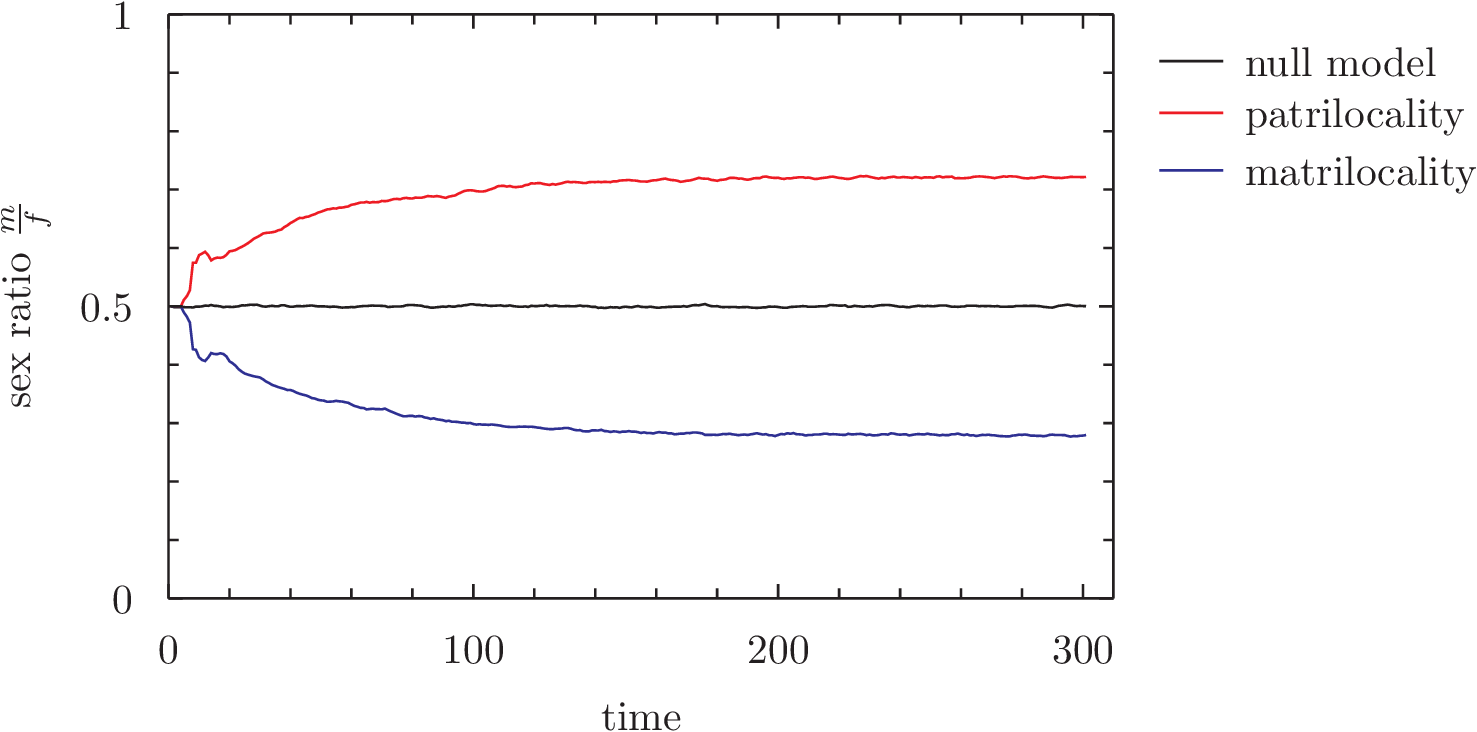
At each time-step, we measure the sex ratio of the population. This is what happens (wordpress is terrible, click for a clearer image):
Average of 500 runs, seeded from 50 family breeding pairs
The sex-ratio of the population is skewed in the direction of the sex that determines family locality – that is to say, patrilocality alone systematically results in the preferential abortion of female offspring, and a higher ratio of males in the population to females. This model will hopefully lend itself to some further work exploring the role of the dowry in maintaining the system by offsetting the costs of giving away offspring, preferential marrying, and how a shift toward “nuclear” family arrangements may have lifted the cost-benefit situations disadvantaging females (and thus making dowry systems redundant).
UPDATE (09/02/12): Here is the data using an amended null model:
Average of 1000 runs, seeded from 50 individuals
The ‘bump’ at around time-step 20 in the first graph noticed by Sean (see comments section) doesn’t appear here; this was an artefact of seeding with identical pairs that breed and die at the same time. Seeding with individuals has smoothed out that curve; staggering the ages of agents would likely smooth it out further. The extra noise in this model means that the skew is less pronounced than before; note the Y axis is zoomed in to ~0.3 – 0.7. The null model here is of the null hypothesis; abortion still happens, the only difference is that instead of a married agent appending to their spouse’s family, the married couple form a new family pair (ie. no matri/patrilocality). This means that any single given population’s sex ratio is susceptible to drift; early, small aberrations toward male or female will become magnified over time. This is, however, equally likely to happen for either male or female, and so the average of 1000 populations shown here is stable at 0.5.
Further implications:
An important additional observation in Brooks’ paper is an examination of the wider social consequences of this particular set of circumstances. The paper names elevated levels of “men competing furiously for wealth and status” as well as “risk-taking, violence, gambling, alcohol and drug abuse, kidnapping and trafficking of women, and the use and abuse of prostitutes” as consequences of surplus males in the population. The implication is that, by this model, these large societal problems can be addressed at least in part by balancing parental investment in children of both sexes, which would be remarkable.
At first blush, the idea that violence results from a surplus of men who don’t have a good enough chance at mating with women has some worrying and problematic implications. It is, nevertheless, intuitively true within a culture of male entitlement, which is something that feminists have long observed – that male violence is the result (and the maintenance) of a patriarchal social order. Since patrilineage, and patrilocality in family structure specifically, are identified as the preconditions for preferential parental investment in males, the eradication of this social order is a necessary step in redressing the sex-ratio balance. The end of patrilineal traditions and patrilocality are also a step toward dismantling a culture of male entitlement more broadly. As a direct consequence, then, this strategy dismantles the structures supporting male entitlement itself at the same time as addressing the skewed sex ratio, and does not simply consist of “giving the men more women to stop them fighting”.
It seems to me that a feminist account that names a culture of male entitlement as the cause for violent female oppression, and an evolutionary account that names structural entitlement systems as the cause for the mass devaluation/infanticide of female offspring are very much on the same page. This approach also very clearly illustrates the compatibility between evolutionary analysis and the socio-economic determinism that is fundamental to radical political thought, precisely by demonstrating how population-wide behaviour can directly result from external economic and social parameters, rather than some innately predisposed condition. We hope that this is at least one small demonstration of how evolutionary psychology and social justice can be rather natural allies.
The Call for Papers can be found here (Deadline for paper & poster submission is September 1, deadline for workshop proposals April , 2013).
To quote from the website:
“The Evolangconference series provides the major meeting for researchers worldwide in the origins and evolution of language. The Evolang conferences are interdisciplinary, with contributions from disciplines including, but not limited to: anthropology, archeology, artificial life, biology, cognitive science, genetics, linguistics, modeling, paleontology, physiology, primatology, and psychology. Typically, about 300 delegates attend, with representatives from all these disciplines. Additional information on Evolang can be found here.”
UPDATE: This paper is now a Trends in Cognitive Sciences Free Featured Article and is available for free here
Noam Chomsky, who infamously stated that the field of language evolution research is “a burgeoning literature, most of which in my view is total nonsense” (see, e.g. here), has a new paper on the topic in press (together with linguist Robert Berwick and neuroscientists Angela Friederici and Johan Bolhuis) called Evolution, brain, and the nature of language (here, unfortunately it’s behind a paywall).
Here’s the abstract:
Language serves as a cornerstone for human cognition, yet much about its evolution remains puzzling. Recent research on this question parallels Darwin’s attempt to explain both the unity of all species and their diversity. What has emerged from this research is that the unified nature of human language arises from a shared, species-specific computational ability. This ability has identifiable correlates in the brain and has remained fixed since the origin of language approximately 100 thousand years ago. Although songbirds share with humans a vocal imitation learning ability, with a similar underlying neural organization, language is uniquely human.
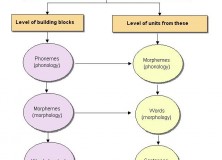
Also interesting is their figure on the Desing of the language system:
“The basic design of language. There are three components: syntactic rules and representations, which, together with lexical items, constitute the basis of the language system, and two interfaces through which mental expressions are connected to the external world (external sensory-motor interface) and to the internal mental world (internal conceptual-intentional interface).”
This still looks very much like the model advocated in for example, the influential and controversial Hauser/Chomsky/Fitch 2002 Science paper (see e.g. here) and from a brief look through the review. The paper also reiterates the view that language is primary an instrument aiding internal thought, and its use for communication is a later by-product (a view that has been thouroughly criticized, by for example Steven Pinker and Ray Jackendoff, e.g. here):
“communication, an element of externalization, is an ancillary aspect of language, not its key function, as maintained by what is perhaps a majority of scholars (cf. [Jim Hurford, Michael Tomasello], among many others). Rather, language serves primarily as an internal ‘instrument of thought’”
Here’s a link to another conference that might be of interest:
The 3rd Linguistic Conference for Doctoral Students will take place at Heidelberg University, Germany from 05.-06. April 2013. The overarching topic of the conference will be: “Interdisciplinary Perspectives on Language, Discourse, and Culture.” The deadline for submissions is 15 February.
I’ve included the Call for Papers below (The Call for Papers can also be downloaded here):
For those of you who might be interested: Language and Cognition has a special issue on the nature and emergence of duality of patterning (paywall access, sorry!). As one of Hockett’s (1960) design features, duality of patterning is the property of human language that enables parts of language to be recombined in a systematic way to create new forms. In the introductory paper, de Boer, Sandler & Kirby (2012) identify two distinct levels where we see duality of patterning: combinatorial (meaningless sounds can be combined into meaningful morphemes and words) and compositional (morphemes and words can be combined to create new constructions with different meanings).For Hockett, not only is duality of patterning a design feature of language (in that all human languages have it), but also it is a unique characteristic of human language.
These two assumptions have been challenged on several fronts. First of all, simple combinatorial structure is found in systems of primate vocalisations, albeit restricted to a relatively limited set of signals. Meanwhile, in the Al-Sayyid Bedouin Sign Language (ABSL), the community does not have a conventionalised level of meaningless elements (although it does have compositional structure at the levels of morphology and syntax). These two examples offer important insights into the duality of patterning debate:
We see then from the case of ABSL that the need to express a large set of signals does not necessarily lead to combinatorial structure, while conversely from the animal systems, it appears that combinatorial structure does not necessarily need a very large set of signals to emerge. As combinatorial structure is the main defining characteristic of duality of patterning, it appears that both the status of duality of patterning as a design feature of language and the evolutionary pathways leading to it need to be rethought. (de Boer, Sandler & Kirby, 2012: 252).
The rest of the special issue is divided up between theoretical and experimental/modelling contributions. The abstracts and links to the papers (again, paywall, sorry!) are posted below. In summary, the general picture emerging from these papers is that duality of patterning is not a clearcut design feature of language, and nor is it necessarily a unique property of our capacity for language. Furthermore, we should also show a greater appreciation of the role that cultural evolution plays:
An apparent point of consensus from the papers in this special issue is that we should not see duality of patterning as a feature hard-wired into an innate language faculty, but rather as arising from multiple pressures operating on language as it emerges and changes in socially interacting populations. When we talk about the evolution of this design of language, then, we are referring more to cultural rather than biological evolution […] It appears that duality of patterning is a rather general state towards which sufficiently complex systems of signals evolve for different reasons: distinctiveness, learnability and a tendency to keep meaningful distinctions, while at the same time trying to make one’s utterances sound similar to those of others in the population. Thus, multiple cognitive processes seem to lead to duality of patterning and therefore, there are probably multiple evolutionary pathways that lead to duality of patterning as well. (de Boer, Sandler & Kirby, 2012: 257).
Sorry for the hyperbole in the title, but now I’ve got your attention – Researchers at the University of Edinburgh have found a gene which is implicated in human brain development which humans have, but chimpanzees don’t.
The study compared the human genome to 11 other species of mammals, including chimpanzees, gorillas, mice and rats, and found that miR-941 is unique to humans.
miR-941 is now being slated as a gene that contributed to how early humans developed tool use and language. This gene, in contrast to the likes of FoxP2, has a very specific function rather than being a gene that down regulates other functions. It is said to be the only gene discovered that has such a specific function while only being present in humans. It is active in two areas of the brain that control our linguistic abilities and also our decision making.
The authors estimate that it emerged between six and one million years ago and that it emerged fully functional out of non-coding genetic material (“junk DNA”) in a very short interval of evolutionary time.
I’m sure the studies where they implant it into mice will start soon. Watch this space.